4. Fattori sensoriali e percettivi nelle funzioni emotive del cervello trino |
|
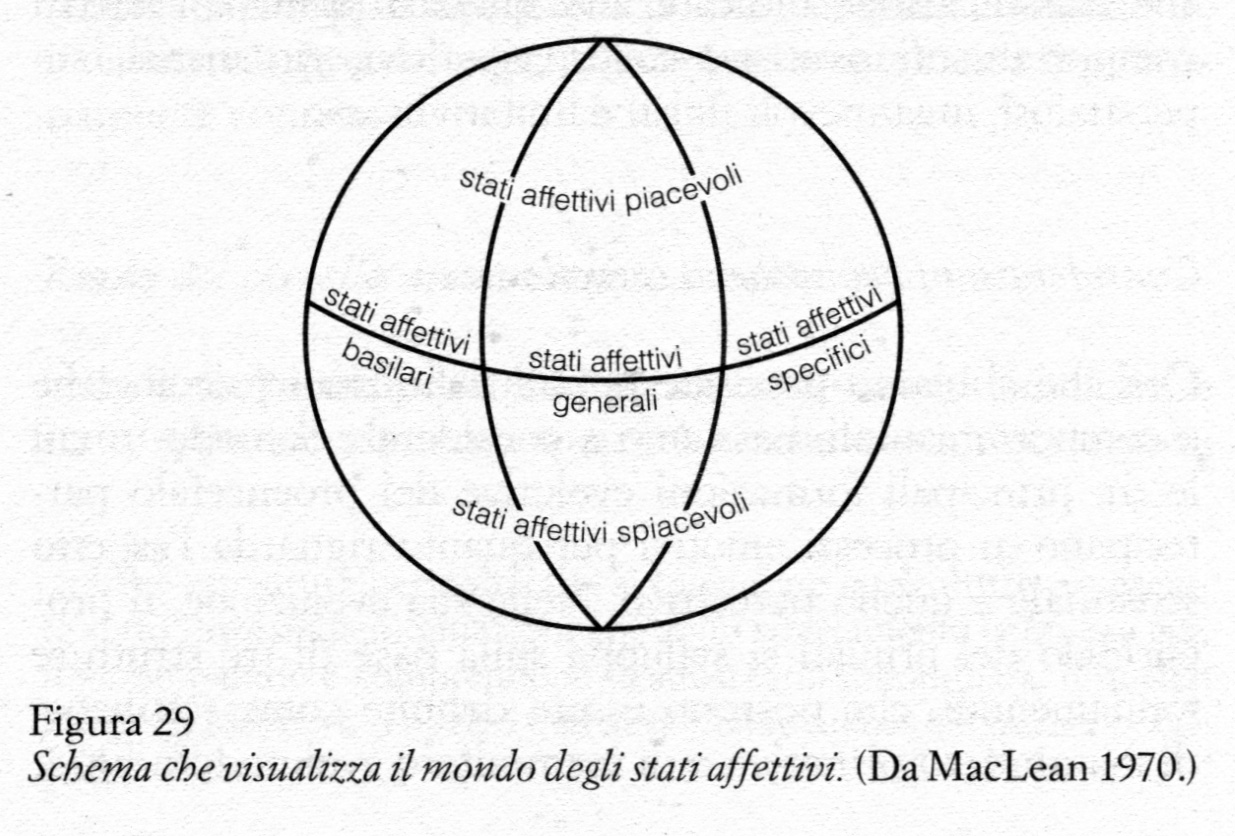
Nel mondo della letteratura e dell'arte ci sono innumerevoli esempi dell'importanza che gli uomini, capaci di introspezione, attribuiscono al ruolo della sensazione e della percezione nel produrre sentimenti emotivi. Si aggiunga a questo ciò che è stato scritto sull'argomento in campi quali la religione, la filosofia, la psicologia e la medicina, e la quantità di informazioni sarà tale da sovraccaricare l'output di un normale calcolatore. Si immagini tuttavia che un calcolatore debba limitarsi a prendere in considerazione i casi in cui le tre parole «sensazione», «percezione» ed «emozione» sono usate per indicare funzioni cerebrali; immediatamente il flusso d'uscita dei riferimenti concreti si ridurrebbe a un esiguo rivoletto. Ma anche questo rivoletto ci fornirebbe informazioni molto più numerose di quante ne potremmo affrontare nei limiti di questo capitolo. Se per esempio dovessimo esaminare le «cause prime», dovremmo necessariamente analizzare l'ampia letteratura sui recettori e passare in rassegna gli effetti della deprivazione sensoriale, in qualunque modo determinata, sull'esperienza emotiva e sulla sua espressione. Per tale ragione, delimitando arbitrariamente l'argomento da trattare, mi occuperò principalmente della questione irrisolta del modo in cui i meccanismi delle sensazioni e delle percezioni esercitano la loro influenza sulle strutture proencefaliche che si ritengono coinvolte nell'emozione. Cercando di affrontare con ordine questo problema, esaminerò una dopo l'altra le principali formazioni evolutive del proencefalo. Procederò, in un certo senso come in uno scavo archeologico: comincerò dalla superficie, esaminando la formazione più recente, per passare poi alle formazioni più antiche. Per ragioni che saranno chiarite più avanti, dedicherò una particolare attenzione al sistema limbico, che dal punto di vista filogenetico rappresenta un'eredità che ci perviene dai mammiferi primitivi. Nella conclusione, poi, richiamerò l'attenzione su alcuni interrogativi spesso trascurati e che probabilmente hanno attinenza con le funzioni del principale equivalente del proencefalo rettiliano che si trova nei mammiferi. Quando scriviamo qualcosa, usiamo le parole sia come materiale per costruire il discorso sia come strumenti per esprimere il pensiero. Dato che non c'è unanimità sul significato dei termini psicologici usati in questo capitolo, sarà opportuno tener presenti le definizioni convenzionali riportate qui di seguito per quanto riguarda l'uso di alcune parole chiave quali «sensazione», «percezione» ed «emozione». Definizioni L'elemento della soggettività è quello che distingue nel modo più chiaro le funzioni cerebrali che sono psicologiche da quelle che non lo sono (MacLean 1960). Perfino i processi cosiddetti inconsci richiedono probabilmente l'esistenza dello stato soggettivo. Il caso del sonno non costituisce un'eccezione, perché l'introspezione rivela che quando si sogna è sempre presente un sentimento di soggettività. La soggettività rimanda alla consapevolezza, associata con varie forme di informazione psicologica. Un filosofo come Kant avrebbe potuto considerarla una «forma di coscienza» a priori. Per parafrasare Spencer (1896), si può dire che la psicologia obiettiva comincia con la psicologia soggettiva. Oltre a ciò che imprecisamente identifichiamo come consapevolezza o coscienza, l'introspezione rivela cinque classi principali di informazione psicologica, che saranno considerate qui di seguito sotto le categorie provvisorie di sensazioni, percezioni, propensioni, emozioni e intellezioni. Tutti questi elementi della psiche di per se stessi non sono altro che informazione. Come Wiener (1948) affermò, più sinteticamente di quanto avessero fatto Berkeley e Hume: «L'informazione è informazione, non è né materia né energia». Nello stesso tempo, è empiricamente dimostrato che non può esserci comunicazione di informazione senza il tramite di ciò che chiamiamo entità di comportamento. L'affermazione di questa invarianza potrebbe essere considerata una legge della comunicazione. L'informazione stessa è considerata un qualcosa di ordinato o, in altre parole, l'ordine che emerge da uno sfondo di disordine. Più grande è il rapporto fra ordine e disordine, più grande è la quantità d'informazione. A tale riguardo, l'informazione astratta si presta a una quantificazione. (Nella teoria dell'informazione, la parola informazione è usata in senso stretto per indicare una quantità numerica che è la misura dell'incertezza in un sistema di comunicazione. Nel contesto presente essa è usata nel suo significato più ampio, per indicare qualsiasi cosa che abbia un senso.) Il modo di procurarsi e di comunicare l'informazione negli animali dipende dalle entità comportamentali del sistema nervoso. Sebbene l'introspezione di per se stessa possa non dare alcuna indicazione su come funzionano queste entità comportamentali (cfr. MacKay 1970), essa è - come è stato già detto - il primo passo che si compie quando si vuole fare un'indagine. Come definiremo le sensazioni e le percezioni? Per cominciare, possiamo dire che la sensazione rappresenta i sentimenti grezzi che, in circostanze normali, dipendono dallo scatenamento di impulsi prodotti dall'attivazione degli enterocettori e degli esterocettori. Per usare la terminologia di Sherrington, le sensazioni rientrano nelle due grandi classi fondamentali delle esterocezioni e delle enterocezioni. Vengono distinte in base alla qualità (modalità) e all'intensità. Le sensazioni, presentandosi da sole o combinate insieme, forniscono una quantità d'informazione maggiore quando vengono valutate in termini di tempo e di spazio. Una volta che hanno ricevuto un tale inquadramento, esse vengono riconosciute introspettivamente come percezioni. Distinzione tra sensazione e percezione da una parte e processo mentale dall'altra. Con l'eccezione di alcune condizioni patologiche, normalmente le sensazioni e le percezioni dipendono da segnali in arrivo al cervello provenienti dai sistemi afferenti specifici e cessano di esistere quando tale attività è terminata. Si confronti questa situazione con quella riguardante le altre tre classi principali di informazione psicologica, e cioè le propensioni, le emozioni e le intellezioni. Esse si distinguono dalle sensazioni e dalle percezioni perché possono verificarsi «dopo il fatto». Il processo inesplicato che rende possibile ciò viene indicato come processo mentale. In termini di attività del sistema nervoso, si potrebbe dire che il processo mentale consiste in una riproduzione neurale - capace di autorigenerarsi - degli eventi così come si sono verificati la prima volta, oppure in qualche nuova disposizione. Rimane un mistero in che modo la disposizione originale degli eventi possa essere conservata (memorizzata) o riprodotta in un nuovo ordine (cioè immaginata o concepita). Dei tre termini psicologici presenti nel titolo di questo capitolo, devo ancora esaminare la definizione di emozione. Si è soliti prendere in considerazione sia l'aspetto soggettivo sia quello espressivo delle emozioni. Io userò l'espressione «stato affettivo» per indicare lo stato soggettivo. Noi possiamo avere esperienza di uno stato affettivo solo in quanto individui. L'esistenza di stati affettivi in un altro individuo deve essere dedotta attraverso qualche forma di comportamento verbale o non verbale. Tale comportamento può essere designato appropriatamente con il termine «emozione», usato nel senso originariamente proposto da Cartesio (1967). Gli stati affettivi differiscono da altre forme di informazione psicologica per il fatto che sono impregnati di una qualità «fisica», che può essere piacevole o spiacevole. Non esistono stati affettivi neutri, perché, parlando dal punto di vista delle emozioni, è impossibile sentire non emotivamente. Nella figura 29 si mostra in che modo gli stati affettivi piacevoli e spiacevoli possono essere suddivisi in tre categorie principali, che ho indicato come stati affettivi basilari, stati affettivi generali e stati affettivi specifici.
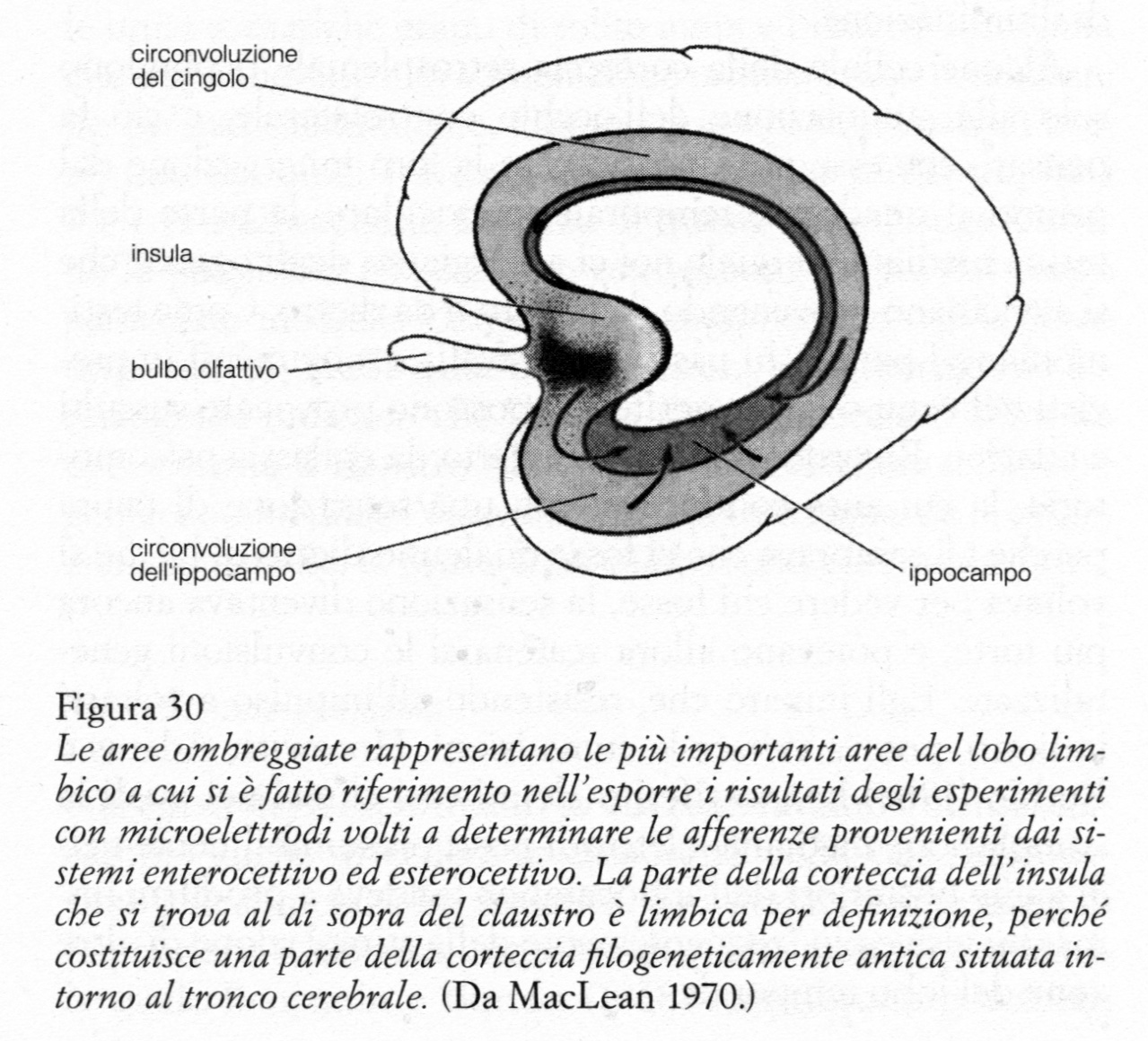
Gli stati affettivi basilari e quelli specifici sono stati affettivi primari, in quanto dipendono in modo diretto rispettivamente dalle enterocezioni e dalle esterocezioni. Gli stati affettivi basilari derivano da enterocezioni che segnalano differenti tipi di stati interni legati a necessità corporee fondamentali, quali il bisogno di cibo, di acqua, di sfogo sessuale, di sonno, e bisogni connessi con i vari organi emuntori. Gli stati affettivi specifici si riferiscono alle esterocezioni e alle percezioni generate direttamente dall'attività di un sistema sensoriale specifico. Alcuni sono innati, mentre altri sono condizionati. Questi ultimi comprendono gli stati affettivi estetici, che si identificano con gli aspetti piacevoli e spiacevoli della musica e delle varie forme di arte. Esempi di stati specifici innati sono quelli suscitati da odori ripugnanti, da rumori allarmanti e da intensi lampi di luce. Gli stati affettivi generali sono secondari, in quanto derivano dagli stati affettivi primari; ma, grazie ai processi mentali ricordati sopra, possono persistere o ripresentarsi «dopo il fatto». Li ho chiamati stati affettivi generali perché possono riguardare sensazioni suscitate da altri individui, da situazioni o da cose. Tutti gli stati affettivi generali possono essere considerati dal punto di vista dell'autoconservazione e della conservazione della specie. Quegli stati affettivi generali che informano circa minacce contro il sé o contro la specie sono spiacevoli, mentre quelli che segnalano l'allontanamento della minaccia o la soddisfazione di un bisogno sono piacevoli. Escludendo il comportamento verbale, ci sono sei tipi principali di comportamento animale e umano che identifichiamo in varia misura con l'esperienza affettiva e con l'espressione emotiva. Si tratta dei comportamenti: 1) di ricerca, 2) di aggressione, 3) di protezione, 4) di depressione, 5) di soddisfazione, 6) di affettuosità. Gli stati affettivi che accompagnano questi comportamenti possono essere descritti in senso lato con i termini: desiderio, rabbia, paura, dolore, gioia, affetto. Il linguaggio simbolico e il processo introspettivo permettono di identificare molte varianti di questi stati affettivi, ma nelle ricerche sugli animali, le deduzioni sugli stati emotivi devono essere basate in larga misura su questi sei tipi generali di comportamento. Ci sono parecchi comportamenti che, in un primo momento, potrebbe sembrare che non si adattino a queste categorie, ma che in un secondo tempo risultano riconducibili alla classificazione indicata. Fra questi i principali sono i comportamenti ossessivo-coatti, ripetitivi, ritualistici, superstiziosi, ingannevoli (falsi) e imitativi. Considerazioni evolutive: il cervello trino Ora che abbiamo precisate queste definizioni psicologiche e comportamentali, passiamo a considerare il modo in cui le tre principali formazioni evolutive del proencefalo partecipano ai processi emotivi per quanto riguarda l'aspetto sensoriale e quello percettivo. Nella sua evoluzione, il proencefalo dei primati si sviluppa sulla base di tre strutture fondamentali, che possono essere definite come rettiliana, di mammifero primitivo e di mammifero evoluto (cfr. fig. 1, p. 194). Ne risulta una notevole compenetrazione fra tre tipi cerebrali che sono radicalmente differenti dal punto di vista chimico e strutturale e che, in senso evolutivo, sono entità separate. Esiste, per così dire, una gerarchia di tre-cervelli- in-uno, ovvero ciò che io chiamo per brevità un cervello trino (cfr. MacLean 1970). Siamo arrivati alla conclusione che ciascun tipo di cervello ha la sua intelligenza specifica, la sua specifica memoria, il suo senso del tempo e dello spazio e il suo tipo particolare di attività e di altre funzioni. Sebbene i tre tipi di cervello abbiano fra loro estese connessioni e dipendano funzionalmente l'uno dall'altro, ci sono elementi che indicano come ciascuno sia in grado di funzionare con una certa indipendenza. Il principale equivalente del proencefalo rettiliano nei mammiferi comprende il corpo striato (nucleo caudato e putamen), il globo pallido e le strutture peripallidali. (Per «strutture peripallidali» si intendono le strutture conosciute sotto diverse denominazioni che sono strettamente connesse con il globo pallido e comprendono la substantia innominata, il nucleo basale di Meynert, il nucleo dell'ansa peduncularis e il nucleo endo-peduncolare.) Il cervello di mammifero primitivo è rappresentato dal sistema limbico, così chiamato in seguito a un mio suggerimento del 1952. La maggior parte della corteccia antica dal punto di vista filogenetico è contenuta nel lobo limbico che circonda il tronco cerebrale (fig. 2, p. 200), ed è conformata un po' come uno stampo del corpo striato che riveste (si veda la fig. 2 in MacLean 1972a). Il cervello di mammifero evoluto è rappresentato dalla neocorteccia, che è in rapida evoluzione, e dalle strutture del proencefalo con le quali essa è primariamente connessa. Ruolo del cervello di mammifero evoluto Già da molto tempo, sulla base di osservazioni chimiche, è stato appurato che aree specifiche della neocorteccia sono connesse rispettivamente ai sistemi somatico, uditivo e visivo, e che queste aree sono essenziali per avere sensazioni e percezioni normali. La derivazione evolutiva di questi sistemi indica che la neocorteccia è orientata primariamente verso l'ambiente esterno. Come ho commentato altrove, «segnali che vengono recepiti da questi tre sistemi sono gli unici che si prestano a essere amplificati elettronicamente e a essere trasmessi via radio» (1972b). Studi anatomici ed elettrofisiologici hanno dimostrato che esiste un sistema ordinato di proiezione che va dai recettori al talamo e dal talamo alla neocorteccia, per cui si stabilisce un collegamento topografico preciso «punto per punto». La tecnica dei potenziali evocati ha reso possibile la suddivisione delle aree somatica, uditiva e visiva in un certo numero di sottoaree; fino a oggi sono state così individuate le aree visive primaria, secondaria e terziaria, le aree uditive primaria, secondaria e terziaria (L'area uditiva V è localizzata al di sopra della corteccia limbica dell'insula) e le aree somatosensoriali primaria, secondaria e supplementare. E’ chiaro che, con l'encefalizzazione, nonostante che il sistema nervoso rimanga ampiamente ridondante, la neocorteccia diventa sempre più la struttura decisiva per la sensazione e per la percezione, e che certamente all'interno della stessa neocorteccia esiste un ordinamento gerarchico. I primati superiori, per esempio, sono ciechi a tutti gli effetti se sono privati della corteccia striata primaria. Ciò è esattamente l'opposto di quanto Snyder e Diamond (1968) hanno recentemente osservato nella tupaia, un animale che si ritiene rappresenti un precursore dei primati. Secondo quanto essi scrivono, «dopo l'asportazione completa del sistema genicolato-striato, la tupaia conserva ancora la visione delle forme e delle strutture e anche la capacità di localizzare visivamente gli oggetti nello spazio». Anche dopo l'asportazione di tutte le aree visive conosciute della neocorteccia, questi animali sono in grado di differenziare righe orizzontali da righe verticali. Il lavoro di Diamond e dei suoi collaboratori (Harting, Hall, Diamond e Martin 1973) ha anche aggiunto nuovi elementi alle conoscenze che si stanno accumulando sul percorso anatomico delle vie sensoriali che raggiungono la neocorteccia diverse da quelle classiche. Essi hanno dimostrato che esiste una via collicolo-pulvinar-temporale, che presumibilmente giustifica la conservazione della funzione visiva nelle tupaie a cui è stata asportata la corteccia striata (Snyder e Diamond 1968). In un articolo del 1963, Myers aveva descritto le proiezioni del collicolo alla porzione inferiore del pulvinar nella scimmia. Parlerò ancora di questa via quando prenderò in esame le connessioni del sistema limbico. Gli studi elettrofisiologici hanno messo in evidenza numerose proprietà funzionali delle aree sensoriali della neocorteccia che si pensa siano coinvolte nella percezione della forma. Per esempio, si ritiene che da questo punto di vista sia di fondamentale importanza il «contorno inibente» scoperto indagando sui campi recettivi di cellule dell'area somato-sensoriale (Mountcastle, Davies e Berman 1957) e dell'area visiva (Hubel e Wiesel 1962). Studi su singole cellule del sistema visivo hanno rivelato che esistono unità che rispondono specificamente ai margini degli oggetti, al contrasto, all'orientamento, al movimento direzionale, al colore, ecc. I dati sperimentali hanno favorito la nascita di un linguaggio magico nel quale si fa riferimento a «neuroni superspecializzati» che sono «riconoscitori di sagome», «riconoscitori di movimento» e così via, quasi possedessero le proprietà soggettive delle monadi di Leibniz e fossero stati creati appositamente per riconoscere solo uno o due tipi di stimoli, mentre in realtà è la rete nervosa alla quale appartengono che può spiegare la loro risposta selettiva. È abbastanza curioso il fatto che, come fa notare Michael (1969), la retina di animali quali la rana, lo scoiattolo di terra e il coniglio, abbia la capacità di discriminare, possieda cioè una funzione che negli animali superiori può essere svolta solo dalla corteccia visiva, in particolare per quanto riguarda l'individuazione di sagome, dell'orientamento e della direzione del movimento. Negli studi neurofisiologici sui meccanismi della percezione, di solito è implicito che il principale scopo perseguito sia quello di capire in che modo gli animali riescono a riconoscere forme ben definite, come se questo aspetto della percezione fosse quello in assoluto più importante per l'organismo. Invece, fino a oggi, non è stato preso sufficientemente in considerazione un problema fondamentale di tipo opposto, e cioè che cosa rende un animale capace di reagire alle configurazioni ambientali, comprese le rappresentazioni parziali e non ben definite di un oggetto o di un animale, le quali provocano un comportamento emotivo di propensione o di ripulsa e che hanno anche, per esprimersi nella terminologia etologica, la funzione di «scatenatori» di forme specifiche di comportamento. Dei molti esempi possibili, il meglio conosciuto è quello del bambino piccolo che risponde alle rappresentazioni schematiche e parziali della faccia umana. Ritornerò su tale questione quando esaminerò le funzioni del sistema limbico e quelle della più importante struttura che costituisce il corrispettivo del proencefalo rettiliano nei mammiferi. Durante le operazioni sul cervello si è appreso che la stimolazione della corteccia somatica, uditiva e visiva può suscitare sensazioni grezze (si veda Penfield e Jasper 1954). Nel tentativo di localizzare i fuochi epilettogeni, Penfield ha trovato che, in alcuni individui, la stimolazione del cervello nelle cosiddette aree associative della corteccia temporo- occipitale può provocare illusioni o allucinazioni uditive e visive (Penfield e Perot 1963). Ma né le osservazioni cliniche né quelle sperimentali hanno consentito di individuare il circuito nervoso mediante il quale l'informazione verbale e non verbale proveniente dai sistemi visivo, uditivo e somatico produce stati affettivi. Fra gli stati affettivi specifici, il dolore è quello che ha ricevuto la maggiore attenzione. Nella loro analisi dei disturbi associati con lesioni talamiche, Head e Holmes (1920) notarono che qualche volta semplici suoni, così come vari stimoli somatici, acuivano il senso del dolore nel lato colpito del corpo. Oltre a ciò, essi descrivevano una strana situazione che, in un certo senso, è l'inverso di quella che stiamo considerando. Secondo loro, nessuno sembra essersi accorto del fatto che stati emotivi possono suscitare sensazioni diverse nelle diverse parti del corpo. «Uno dei nostri pazienti» riferivano fra l'altro «non era in grado di andare al suo luogo di culto perché, come diceva, "non riusciva a sopportare gli inni sacri nel lato colpito del corpo".» Sulla base di casi del genere, essi giunsero alla conclusione che il talamo ottico era la fonte delle emozioni. Considerazioni introduttive allo studio delle formazioni limbica e rettiliana Von Economo (1931) scoprì alcune zone più ventrali che avevano un'importanza fondamentale per l'esperienza emotiva. Nella sua monografia Encephalitis Lethargica, egli arrivò alla conclusione che il tegmento, la parete basale e quella posteriore del terzo ventricolo e la regione dell'acquedotto erano il «bersaglio preferito» della malattia. Egli sottolineò come questi dati sperimentali fossero coerenti con l'osservazione che fra i sintomi persistenti «la difficoltà di suscitare emozioni... è soprattutto un difetto primario». Egli indicò che alcuni pazienti possono aver fame, ma non ricevere piacere dal mangiare, o riconoscere la sensazione del freddo senza provare il normale senso di freddo. In generale, essi possono lamentarsi di «sentirsi come un vulcano esaurito». C'è da osservare che le strutture di cui in particolare parla von Economo si trovano nella regione irrorata dai vasi sanguigni perforanti della fossa interpeduncolare (cfr. Mett- ler 1955). Grazie a recenti indagini, che si possono in parte far risalire a Ulf S. von Euler, sappiamo oggi che in questa regione esistono neuroni contenenti dopamina, i quali innervano il corpo striato, compreso il cosiddetto striato olfattivo. Vista la profonda influenza esercitata da questo sistema ascendente sul comportamento spontaneo degli animali, ci possiamo chiedere retrospettivamente fino a qual punto l'ottundimento emotivo messo in evidenza da von Economo era dovuto alla distruzione delle cellule dopaminergiche della sostanza nera e di quelle della rete nervosa che si estende attraverso tutta la fossa interpeduncolare (gruppi A9 e AIO di Dahlström e Fuxe 1964). Mi si consenta di riferire due casi (non pubblicati) che illustrano il problema. Abbiamo trovato che la distruzione di questi due gruppi di cellule mediante la idrossi-dopammina nella scimmia scoiattolo provoca, in un primo momento, la catalessi, seguita poi da un quadro di parkinsonismo. Sintomi di catalessi sono riconoscibili già prima che la scimmia si riprenda completamente dall'anestesia. Retrospettivamente, si potrebbe pensare che la catalessi osservata da Ingram, Barris e Ranson (1936) nei gatti in seguito a lesioni localizzate fra i corpi mammillari e il terzo nervo fosse una conseguenza del danneggiamento di questo stesso gruppo di cellule. All'inabilità provocata dalle lesioni in questa regione si può contrapporre l'effetto mobilitante dell'apomorfina, la quale agisce, a quanto pare, sui recettori della dopammina. Uno studio comparato ha rivelato che l'apomorfìna provoca un aumento di attività non finalizzata in animali delle specie più diverse, come pappagalli, tacchini, opossum e scimmie scoiattolo. Un tacchino, per esempio, può correre senza uno scopo per tre o quattro ore avvicinandosi e poi allontanandosi dal gruppo degli altri tacchini. Le due opposte condizioni che ho descritto potrebbero servire a rafforzare le altre prove del fatto che i sistemi ascendenti dopamminergici esercitano un'influenza «attivizzante» sul comportamento animale. Estrapolando dagli esperimenti sugli animali di Ranson (1939) e di altri, si potrebbe andare più avanti di von Economo e dire che le più importanti vie maestre che portano all'espressione della personalità di base passano attraverso il diencefalo ventrale e che i fasci laterale e mediale del proencefalo sono i più importanti sistemi di fibre che assicurano rispettivamente il collegamento con le strutture striate e limbiche, sia in senso centrifugo che in senso centripeto. Il fascio laterale del proencefalo comprende l'ansa e il fascicolo lenticolare, come anche le vie strio-nigra e nigro-striata. L'ansa lenticolare, quando emerge dalla parte rostro-mediale del globo pallido, ha l'aspetto di un fusto di coda di cavallo (equiseto), con le sue componenti compatte e quelle diffuse che scorrono attraverso la parte dorso-laterale dell'ipotalamo e che in parte si intrecciano con il fascio mediale del proencefalo. Rileggendo la letteratura sull'argomento, è interessante vedere come i ricercatori abbiano dato scarsa importanza o abbiano del tutto trascurato il significato delle parti compatte e di quelle ampiamente diffuse dell'ansa, e lo stesso si può dire per il fascicolo lenticolare. Può darsi che ciò sia dovuto in parte alle conclusioni di Ranson (1939) secondo cui queste connessioni dello striato non dovevano avere una funzione importante per le alterazioni emotive osservate nelle scimmie rhesus con lesioni bilaterali dell'ipotalamo laterale. Eppure questo autore affermò nello stesso tempo che le lesioni dell'ansa provocavano una completa scomparsa dei neuroni nel segmento mediale del globo pallido. Si potrebbe notare, per inciso, che l'ansa non è semplicemente una via pallidofuga. Oggi è chiaro che, analogamente al fascio mediale del proencefalo, essa contiene fibre ascendenti (Carpenter e Peter 1972). Jacobowitz e io (lavoro non pubblicato) abbiamo trovato, sperimentando sulle scimmie scoiattolo, che in essa sono presenti fibre ascendenti dopaminergiche, alcune delle quali attraversano l'ipotalamo proprio a lato del fornice. Dati sperimentali ottenuti negli animali dimostrano il ruolo fondamentale delle vie lenticolari per l'espressione del «carattere» di un animale. Nel corso delle ricerche sulle scimmie scoiattolo, volte a chiarire i meccanismi cerebrali responsabili del comportamento tipico della specie (si veda più avanti), ho avuto occasione di produrre nella parte ventrale del diencefalo ampie lesioni bilaterali che coinvolgevano il sistema centrale dell'ansa e parte del fascio mediale del proencefalo. Grazie a un'assistenza accurata, molte scimmie sopravvivevano al periodo critico post-operatorio. Esse mostravano un pieno recupero della capacità di camminare e di alimentarsi da sole. Tuttavia, ciò che colpisce in queste scimmie è la completa assenza di ciò che potremmo chiamare la loro animalità. Esse hanno, per così dire, un comportamento da idiota, che è penoso da osservare. Il sistema limbico Una volta poste queste basi generali, possiamo occuparci del sistema limbico, che rappresenta un'eredità che ci proviene dai mammiferi primitivi. Le prove più evidenti della funzione svolta dal sistema limbico nel comportamento emotivo derivano dalle osservazioni cliniche. Le scariche dei neuroni nella corteccia limbica del lobo temporale o nelle sue vicinanze possono scatenare un ampio spettro di vividi sentimenti affettivi. Gli stati affettivi basilari e generali sono di solito del tipo che è in rapporto a minacce contro l'autoconservazione (MacLean 1958a). Più raramente si può trattare di stati affettivi di natura piacevole o estetica, che forse riflettono l'estendersi della scarica convulsiva ad altre regioni del sistema limbico. Gli stati affettivi basilari comprendono quelli della fame, della sete, della nausea, e sensazioni associate con le funzioni emuntorie. Gli stati affettivi specifici comprendono sapori e odori sgradevoli e sensazioni somatiche quali il dolore e il prurito. Fra gli stati affettivi generali ci sono i sentimenti di paura, di terrore, di tristezza, il desiderio di solitudine, la dimestichezza, l'estraneità e, molto raramente, la rabbia. Il sentimento di paura è comunemente riferito alla regione epigastrica e può dare l'impressione che il cuore salga in gola. Come ho messo in rilievo altrove (1952), gli stati affettivi generali sono di solito «fluttuanti», fin tanto che non vengono identificati con una persona o una situazione particolare. Una delle esperienze affettive più comuni è quella cosiddetta del déjà vu. E significativo che, come hanno notato Penfield ed Erickson (1941), il paziente possa avere esperienza solo del sentimento che accompagna l'atto del ricordare. Una situazione analoga si verifica nelle aure, nelle quali si presentano sentimenti del tipo «eureka», espressi da frasi del tipo: «È proprio così, è assolutamente vero!», oppure: «E così che è fatto il mondo!» (MacLean 1970, 1973b). Paradossalmente sembrerebbe che l'antico sistema limbico ci procuri forti e fluttuanti sentimenti affettivi di convinzione, che noi colleghiamo con rivelazioni e con credenze, senza badare al fatto che queste siano vere o false! Casi clinici di epilessia limbica indicano anch'essi che 1) il sistema limbico è fondamentale per sentire affettivamente la realtà di se stessi e dell'ambiente circostante (MacLean 1972b); e che 2) la distruzione in seguito a ictus delle sue funzioni può produrre cambiamenti di umore, distorsioni della percezione, sentimenti di depersonalizzazione, allucinazioni e manie paranoidi (cfr. sopra). Gli aspetti affettivi delle esperienze personali - esemplificati dal déjà vu - sembrano essere un elemento importante per la memoria (MacLean 1969a). Una delle conseguenze delle convulsioni limbiche è l'amnesia che, dal punto di vista temporale, è in rapporto con il momento terminale dell'aura e con l'inizio e la durata dell'automatismo. In qualche caso quest'ultimo comprende attività che quasi certamente richiedono una neocorteccia funzionante. Le sensazioni suscitate dai fuochi epilettogeni possono coinvolgere qualcuno dei sistemi sensoriali. Può trattarsi di sensazioni olfattive, gustative, viscerali e genitali. I suoni possono sembrare insolitamente forti o fievoli; si può avere l'impressione che alcune parti del corpo siano enormemente gonfiate; può verificarsi la condizione di micropsia o di macropsia, nella quale gli oggetti appaiono insolitamente piccoli o grandi. Sensazioni, percezioni, stati affettivi: non esiste alcuna entità clinica che, come l'epilessia limbica, combini insieme nella sua sintomatologia questi tre aspetti psicologici che sono l'oggetto della nostra trattazione. Qual è la base neurale di questa e delle altre manifestazioni a cui abbiamo accennato? Nel mio articolo del 1949 sul «cervello viscerale», io rielaborai la teoria dell'emozione di Papez (1937), avanzando l'ipotesi che gli impulsi provenienti da tutti i sistemi enterocettivi ed esterocettivi arrivino all'ipotalamo attraverso la circonvoluzione dell'ippocampo. Questa ipotetica via di connessione era indicata schematicamente nella figura 3 dell'articolo. La formazione dell'ippocampo era presentata come un meccanismo che combina l'informazione di origine interna e quella di origine esterna in sentimenti affettivi, i quali vengono poi ulteriormente elaborati e trovano la loro espressione mediante le connessioni con l'amigdala, il setto, i gangli basali, l'ipotalamo e il circuito che permette una connessione di ritorno con il lobo limbico, e che oggi è conosciuto come «circuito di Papez». Prima di discutere la questione delle afferenze alla corteccia limbica, si devono far presenti due altre importanti considerazioni. Prima di tutto, gli studi di patologia fatti da Sano e Malamud (1953) e da Margerison e Corsellis (1966) hanno rivelato che la sclerosi del corno d'Ammone è, secondo le parole di Malamud (1966), il «comune denominatore» dei casi di epilessia psicomotoria. Dato che spesso la sclerosi si estende ad altre strutture temporali mediali, Falconer, Serafetinides e Corsellis (1964) preferiscono usare l'espressione «sclerosi temporale mediale». L'ipotesi che tale sclerosi o altre lesioni temporali mediali siano responsabili dei fuochi epilettogeni trova un forte sostegno in due serie di cento casi di epilessia, nei quali Falconer (1970) asportò in un solo blocco il lobo temporale interessato. Questa operazione non solo ha l'effetto di eliminare totalmente o in massima parte il tessuto lesionato, ma consente anche un completo esame anatomo-patologico. In secondo luogo, è da mettere in evidenza il fatto che le scariche convulsive che si originano nella corteccia limbica o nelle sue vicinanze tendono a diffondersi ampiamente tutt'intorno, rimanendo tuttavia confinate entro il sistema limbico. E’ probabile che l'ippocampo sia quasi sempre coinvolto, sia che le scariche convulsive abbiano origine in esso sia che vi si propaghino, provenendo da strutture che sono in rapporto con esso. Registrazioni simultanee dalla neocorteccia possono mostrare che nel corso di tali scariche convulsive si verificano in essa scarse modificazioni, a eccezione di una desincronizzazione generalizzata. Per tale ragione io ho parlato (MacLean 1954) di potenziale «schizofisiologia» dei sistemi limbico e neocorticale e ho ipotizzato che questa situazione può spiegare in parte i conflitti fra ciò che «sentiamo» affettivamente e ciò che «conosciamo». Non è affatto chiaro in che modo le scariche convulsive limbiche possano generare vari fenomeni sensoriali e percettivi, se si fa eccezione per le sensazioni olfattive, rappresentate nel lobo piriforme, e per le sensazioni gustative e viscerali, la cui rappresentazione nella parte limbica dell'insula è tuttavia meno sicura. Ci sono dati sperimentali che dimostrano come le convulsioni limbiche non modificano in modo apprezzabile l'attività bioelettrica delle aree sensoriali primarie della neocorteccia. Gli stimoli acustici, per esempio, continuano a essere efficaci nell'evocare potenziali nell'area uditiva, mentre è in corso una scarica convulsiva che si propaga nell'ippocampo (Flynn, MacLean e Kim 1961; Prichard e Glaser 1966). Nell'articolo del 1949 citato prima, io avanzai l'ipotesi che l'informazione somatica, uditiva e visiva fosse convogliata verso la circonvoluzione dell'ippocampo mediante vie di connessione cortico-corticali provenienti dalle aree primarie di afferenza. Successivamente Pribram e io riferimmo i dati ottenuti nel gatto (MacLean e Pribram 1953) e nella scimmia (Pribram e MacLean 1953) registrando l'attività neuronica dopo trattamento con stricnina; questi dati risultavano compatibili con la mia ipotesi. Più tardi, Jones e Powell (1970) descrissero uno studio anatomico sperimentale nel macaco che rivelava la possibilità di connessioni corticali scaglionate che collegavano queste aree non solo con la circonvoluzione dell'ippocampo, ma anche con la corteccia limbica della circonvoluzione anteriore del cingolo e dell'area orbitale posteriore (si veda in proposito anche lo studio anatomico di Van Hoesen, Pandya e Butters 1972). Dopo una breve ricapitolazione dei risultati ottenuti in passato negli studi elettrofisiologici, riassumerò alcuni esperimenti condotti con la tecnica dei microelettrodi che, sulla base della latenza della risposta, indicano come l'informazione visiva, uditiva, somatica e viscerale raggiunga diverse specifiche zone del lobo limbico attraverso vie subcorticali abbastanza dirette. Allo stesso tempo, riferirò i dati anatomici che confermano queste conclusioni. In uno studio pubblicato nel 1952 (MacLean, Horwitz e Robinson 1952), trovammo che stimoli gustativi e stimoli somatici nocivi avevano l'effetto di evocare nell'area piriforme potenziali simili ai potenziali olfattivi, che si ripetevano ritmicamente. In qualche caso, in seguito a stimolazione olfattiva o gustativa, i potenziali ritmici comparivano nell'ippocampo. Proseguendo questa linea di ricerca, Green e Ar- duini (1954) trovarono che varie forme di stimolazione sensoriale evocavano un'attività theta ritmica nell'ippocampo degli animali non anestetizzati. Effetti simili furono osservati in animali macrosmatici, ma non nei primati, e vennero considerati di natura non specifica. Continuando la ricerca sulle afferenze limbiche, noi usammo microelettrodi per registrare le risposte evocate nei singoli neuroni; gli esperimenti erano fatti su scimmie scoiattolo sveglie e sedute, con impianto cronico dei microelettrodi. Lavorare in queste condizioni ha il vantaggio di evitare gli effetti depressivi dell'anestesia sulla trasmissione neurale e, differentemente da quanto succede quando si usa la tecnica dei macroelettrodi per registrare i potenziali lenti evocati, consente di essere sicuri della localizzazione della risposta neuronica. Fino a oggi abbiamo sottoposto alla prova più di 7500 unità cerebrali, delle quali circa 2500 (33 per cento) erano localizzate nella corteccia limbica. Abbiamo esplorato tutta la corteccia del lobo limbico, a eccezione dell'area piriforme e dell'area orbitale posteriore. Riassumerò ora brevemente i dati già pubblicati ottenuti con la stimolazione visiva, uditiva e somatica. Stimolazione visiva Praticamente tutte le unità capaci di rispondere a stimoli visivi erano localizzate nella circonvoluzione posteriore dell'ippocampo (si veda la fig. 30), nella corteccia paraippocampale della circonvoluzione lingulare e nella corteccia retrospleniale, nelle vicinanze della zona in cui essa confina con la corteccia striata (MacLean, Yokota e Kinnard 1968). Le condizioni dell'esperimento non consentivano di tracciare la mappa del campo recettivo; tutto quello che si poteva fare era di esaminare le cellule un po' qua e un po' là, seguendo uno schema appropriato. La regolarità e il carattere delle risposte agli stimoli visivi, così come i valori di latenza, facevano pensare che le vie di connessione fossero subcorticali piuttosto che transcorticali. A sostegno di questa ipotesi sono i risultati di uno studio neuroanatomico, nel quale vennero usate tecniche raffinate - capaci di mettere in evidenza fibre sottili - per seguire le tracce della degenerazione a partire da lesioni localizzate nel corpo genicolato laterale e nel pulvinar (MacLean e Creswell 1970). Lesioni nella parte ventro-laterale del nucleo genicolato laterale provocavano una degenerazione in quella parte della radiazione ottica conosciuta come anello di Meyer, che fa una deviazione nel temporale e penetra proprio nel centro della circonvoluzione posteriore dell'ippocampo. Fu possibile seguire le tracce di alcune fibre fino alla zona della corteccia limbica capace di rispondere agli stimoli visivi e alle attigue aree neocorticali. Un tipo più grossolano di degenerazione fu rilevato nella circonvoluzione posteriore dell'ippocampo e nelle aree contigue in seguito a lesioni nella parte inferiore del pulvinar. Le proiezioni del pluvinar sono contenute in un fascio di fibre che scorre proprio a lato delle radiazioni ottiche. Questa osservazione richiama alla mente la discussione sulle connessioni, descritte recentemente, che collegano il collicolo superiore con la parte inferiore del pulvinar. Circa metà delle cellule della circonvoluzione posteriore dell'ippocampo dette una risposta positiva prolungata all'illuminazione oculare; ci possiamo allora chiedere se esse possono avere una funzione negli stati di veglia, di vigilanza e di attenzione, oppure nella regolazione delle funzioni neuroendocrine influenzate da cambiamenti diurni e stagionali di illuminazione. Alcune cellule della corteccia retrospleniale rispondono solo alla stimolazione dell'occhio controlaterale, e ciò fa pensare che esse possano ricevere la loro innervazione dal primitivo quadrante temporale monoculare, la parte della retina mediante il quale noi ci accorgiamo degli oggetti che si avvicinano provenendo da un lato o da dietro. Come testimoniano i paraocchi usati per i cavalli, i movimenti imprevisti nel campo visivo periferico possono provocare sussulti e allarme. Ricordo un paziente affetto da epilessia psicomotoria, la cui aura cominciava con una sensazione di paura perché gli sembrava che ci fosse qualcuno dietro di lui. Se si voltava per vedere chi fosse, la sensazione diventava ancora più forte, e potevano allora scatenarsi le convulsioni generalizzate. Egli imparò che, resistendo all'impulso a voltarsi indietro, poteva evitare le convulsioni. Horowitz, Adams e Rutkin (1968) hanno riferito alcuni casi clinici nei quali la stimolazione mediante elettrodi posti presumibilmente nelle «aree posteriori dell'ippocampo» tendeva a produrre immagini visive più frequentemente della stimolazione di altre zone del lobo temporale. La parte della corteccia dell'insula che ricopre il claustro (fig. 30) è limbica per definizione, perché costituisce parte della corteccia filogeneticamente antica che circonda il tronco cerebrale.
Noi abbiamo trovato alcune unità nella corteccia limbica dell'insula capaci di rispondere con una scarica vivace a un oggetto che si avvicina. Questo risultato richiama alla mente le osservazioni di Penfield e Jasper (1954), secondo cui le scariche nella regione parainsulare possono provocare macropsia, una condizione nella quale si ha l'impressione che gli oggetti si ingrandiscano. Stimolazione somatica e uditiva Risposte alla stimolazione somatica e a quella uditiva sono state ottenute solo nella corteccia limbica dell'insula (Suda- kov, MacLean, Reeves e Marino 1971). I campi recettivi delle unità somatiche erano di solito ampi e bilaterali. Le unità che rispondevano alla stimolazione uditiva erano localizzate un po' più caudalmente. C'erano due tipi principali di unità, uno dei quali scaricava con una latenza più corta, fra i 7 e i 15 millisecondi. Latenze di questo ordine suggeriscono la possibilità che esistano connessioni dirette dal corpo genicolato mediale. Come è stato argomentato nell'articolo originale, gli studi sulla degenerazione sia anterograda sia retrograda indicano che la parte della corteccia dell'insula che sovrasta il claustro riceve fibre di proiezione dal corpo genicolato mediale. Viceversa, per quanto riguarda le proiezioni somatiche, le prove anatomiche sono contraddittorie. Stimolazione gustativa Nel corso degli esperimenti in cui esploravamo l'insula, cercammo anche di vedere se esistevano unità che rispondevano alla stimolazione gustativa. Trovammo unità capaci di rispondervi nella zona anteriore di quella stessa regione nella quale Benjamin e Burton (1968) registrarono potenziali evocati in seguito alla stimolazione della corda del timpano nelle scimmie scoiattolo. Altre unità della stessa regione erano attivate dalla stimolazione meccanica della cavità orale, compresa la faringe. Stimolazione del vago Ci è sembrato abbastanza strano constatare che le unità della corteccia cingolata (si veda la fig. 30) siano praticamente incapaci di rispondere alla stimolazione visiva, uditiva e somatica (Bachman e MacLean 1971). In un successivo tentativo di scoprire la natura delle afferenze alla corteccia cingolata, abbiamo cercato di capire se esistevano proiezioni viscerali e, a questo scopo, abbiamo studiato gli effetti di scariche vagali sull'attività dei neuroni. Questi esperimenti sono stati effettuati su scimmie scoiattolo sveglie e sedute, nelle quali in precedenza erano stati impiantati cronicamente gli elettrodi sul vago a livello cervicale o nel punto in cui il vago penetra nel foro giugulare (Bachman, Katz e MacLean 1972b). Sulle oltre 300 unità esaminate fino a oggi, hanno risposto un po' più del 20 per cento, con un rapporto fra unità inizialmente eccitate e unità inizialmente inibite che si aggirava fra 2 e 3. La maggior parte delle unità capaci di rispondere sono state localizzate nella regione centrale del giro. La latenza delle risposte delle unità eccitate era di soli 15 millisecondi, e ciò fa pensare a una via di connessione abbastanza diretta. In uno studio parallelo, abbiamo cercato di imitare la stimolazione naturale dei recettori viscerali, iniettando microquantità di 5-idrossi-triptammina (serotonina) attraverso un catetere introdotto nella vena cava superiore (Bachman, Katz e MacLean 1972a). Delle 82 unità della corteccia cingolata situata sopra al corpo calloso esaminate, il 18 per cento reagirono, e di queste i due terzi mostrarono un aumento della frequenza delle scariche, mentre nel restante terzo ci fu una diminuzione di questa frequenza. Anche queste unità erano localizzate nella parte centrale della circonvoluzione. Gli studi anatomici di Morest (1961) indicano che potrebbe esistere una via ascendente che andrebbe dal nucleo solitario al nucleo tegmentale dorsale di Gudden, dal quale gli impulsi potrebbero salire, attraverso il sistema del peduncolo mammillare, al nucleo ventrale anteriore del talamo e alla corteccia cingolata. I risultati degli studi di istofluorescenza (si vedano, per esempio, Fuxe 1965a; Jacobowitz e Kostrzewa 1971; Olson e Fuxe 1972) non sono incompatibili con l'interessante possibilità che esista un sistema ascendente adrenergico che parte dal nucleo solitario. Il nucleo antero-ventrale del talamo ha numerose terminazioni sottili adrenergiche, e analoghe terminazioni si trovano anche nella circonvoluzione del cingolo. In uno studio comparato di istofluorescenza condotto su vari animali, fra cui l'uistitì pigmeo e la scimmia scoiattolo, Jacobowitz e io (lavoro non pubblicato) trovammo che, per tutti i sistemi amminergici conosciuti, le strutture in cui questi erano organizzati erano state conservate con notevole costanza attraverso l'evoluzione dei primati. Fra le altre aree limbiche che ci proponiamo di esplorare nella scimmia per individuare i neuroni che rispondono alla stimolazione del vago, ci sono l'area orbitale posteriore e l'area anteriore dell'insula, che corrispondono a una regione del cervello del gatto dalla quale numerosi ricercatori hanno registrato alterazioni dell'attività spontanea (Bailey e Bremer 1938), oppure dei potenziali evocati (Dell e Olson 1951a, 1951b; Korn, Wendt e Albe-Fessard 1966), in seguito a stimolazione vagale. Integrazione sensoriale Prima di addentrarci nel problema dell'integrazione sensoriale, vorrei far rilevare il fatto che nei nostri esperimenti nessuna unità dell'ipotalamo rispondeva alla stimolazione visiva o somatica e che solo alcune di esse erano influenzate dalla stimolazione uditiva (Poletti, Kinnard e MacLean 1973). Ciò farebbe pensare che l'informazione sensoriale che agisce sull'ipotalamo viene precedentemente integrata e rielaborata in strutture a esso connesse, come la corteccia limbica. Nei nostri studi iniziali sulle scimmie anestetizzate con alfacloralosio, trovammo alcune unità corticali limbiche che mostravano di avere afferenze sensoriali convergenti. E tuttavia significativo il fatto che nelle scimmie sveglie e sedute tutte le unità capaci di rispondere sembrano essere specifiche per una sola modalità, rivelando dunque un alto grado di selettività. Sorge allora l'ulteriore problema: in che modo l'informazione che dal sistema enterocettivo e da quello esterocettivo raggiunge la corteccia limbica viene integrata e rielaborata? O, esprimendosi in modo metaforico, in quale punto del sistema limbico risiedono gli «osservatori»? Uno dei luoghi probabili è la corteccia entorinale della circonvoluzione dell'ippocampo, la quale riceve vie di connessione dalla corteccia fronto-temporale (aree orbitale posteriore, insulare anteriore, piriforme e temporale polare) e dalla corteccia paraippocampale caudale (fig. 30). La corteccia entorinale occupa nell'uomo un'area estesa; da questa area, l'informazione potrebbe essere trasferita avanti fino all'ippocampo attraverso le vie perforante e alveolare (fig. 4). La parte anteriore dell'ippocampo riceve anche connessioni dalla regione fronto-temporale, ed esistono afferenze alla regione posteriore dell'ippocampo che provengono dalla circonvoluzione del cingolo (attraverso il cingolo), e anche dalla lamina midollare superficiale. Quest'ultima via potrebbe trasmettere l'informazione di provenienza vagale (si veda sopra). Nell'ippocampo il sistema collaterale di Schaffer potrebbe costituire la struttura che permette di mettere in rapporto reciproco le varie informazioni provenienti dalle diverse modalità sensoriali. Il setto, al quale giungono vie di connessione dall'ippocampo, è un'altra possibile fonte dell'informazione entero- cettiva che raggiunge l'ippocampo. Si ritiene che le proiezioni del setto terminino nello stratum oriens, forse sui dendriti basali delle cellule piramidali dell'ippocampo, mentre la via perforante termina sui dendriti apicali. In uno studio a livello intracellulare dei neuroni dell'ippocampo condotto sulla scimmia sveglia e seduta, abbiamo trovato che gli stimoli del setto provocavano la comparsa di potenziali eccitatori postsinaptici associati con scariche neuroniche, mentre la stimolazione del bulbo olfattivo generava potenziali eccitatori postsinaptici, ma mai potenziali d'azione (Yokota, Reeves e MacLean 1970). Per usare la terminologia del condizionamento classico, gli impulsi provenienti da queste due afferenze esterocettive potrebbero essere paragonate rispettivamente a stimoli incondizionati e a stimoli condizionati. Brazier (1964) ha riferito su potenziali lenti evocati da stimoli visivi nell'ippocampo di pazienti che erano sottoposti a test diagnostici per l'epilessia. Efferenze limbiche Non rientra nell'argomento di questo saggio esaminare i meccanismi d'uscita del sistema limbico; ma ci sono alcuni punti salienti che meritano di essere ricordati. Gli studi fatti con la tecnica dei microelettrodi hanno mostrato che le scariche a raffica del fornice o le scariche postume dell'ippocampo inibiscono le risposte dei singoli neuroni della regione intralaminare caudale agli stimoli potenzialmente nocivi del V nervo (Yokota e MacLean 1968). In un articolo successivo (Poletti, Kinnard e MacLean 1973), abbiamo riferito che le scariche a raffica dell'ippocampo provocano risposte in un'ampia percentuale di unità in determinate strutture del proencefalo basale, della regione preottica e dell'ipotalamo. In ognuna di queste strutture, più dell'80 per cento delle unità capaci di rispondere mostrarono un'eccitazione iniziale. Anche le scariche postume dell'ippocampo avevano più frequentemente un effetto eccitatorio sulle unità che un effetto inibitorio. In seguito alle scariche postume, si verificava un'alterazione nella configurazione delle scariche delle unità che duravano da uno a undici minuti. Questi ultimi risultati possono aiutare a spiegare le prolungate modificazioni «di ritorno» nel comportamento e nel sistema vegetativo che sono state osservate dopo le scariche postume dell'ippocampo e che comprendevano da una parte stati di agitazione, e dall'altra un'accentuazione del piacere e delle risposte sessuali. Come già appariva evidente dai dati elettrofisiologici, un parallelo studio neuroanatomico mostrò per la prima volta nei primati che il fornice proietta all'area mediale preottica e alla regione attigua al fornice stesso (Poletti, Kinnard e MacLean 1973). Lo studio di Hess e Briigger (1943) indicava la regione attigua al fornice (la cosiddetta zona intermedia di Hess) come una struttura coinvolta nell'espressione del comportamento di rabbia. L'area preottica mediale è diventata sempre più interessante non solo perché partecipa al controllo dell'attività gonadotropica e della funzione genitale, ma anche a causa del ruolo che svolge nella differenziazione sessuale in alcuni animali macrosmatici. E’ possibile inoltre seguire il percorso di alcune fibre fino alla regione tuberale, in cui lo studio elettrofisiologico ha messo in evidenza una risposta cellulare alle scariche a raffica dell'ippocampo. Tenendo presenti questi risultati e altri analoghi, si dovrebbe prestare attenzione alle prove sempre più numerose secondo le quali la stimolazione dell'ippocampo può avere - in dipendenza dallo stato fisiologico del momento - un effetto facilitante o inibente sulla liberazione dell'Acth, sui riflessi cardiovascolari e sulla reattività viscerale (per i riferimenti bibliografici sull'argomento, si veda Poletti, Kinnard e MacLean 1973). I risultati dei nostri studi mediante microelettrodi indicherebbero che nella scimmia sveglia e seduta siamo ancora molto lontani dal poter attribuire all'ippocampo una gamma di funzioni inibitorie, eccitatorie e regolatrici. Successivamente, Poletti, Sujatanond e Sweet (1972) hanno mostrato che dopo la sezione del fornice le scariche a raffica dell'ippocampo erano ancora efficaci nel suscitare le risposte delle unità nelle strutture esaminate nello studio precedente, ma in questo caso la latenza era più lunga. Sembra probabile che gli impulsi siano trasmessi attraverso l'amigdala, la quale rappresenta una delle più importanti vie maestre per le proiezioni della regione fronto-temporale al sistema limbico. Dal punto di vista clinico, così come dal punto di vista sperimentale per quanto riguarda gli animali, risulta che la formazione dell'ippocampo sia coinvolta nel sogno e in altre manifestazioni del sonno Rem. Data la forte componente affettiva presente nel sogno, questo è un punto che interessa l'argomento che stiamo trattando. Negli ultimi tempi si è rivolta una particolare attenzione a questo fatto, perché si è scoperto un legame fra i sistemi amminergici e i meccanismi del sonno (per una rassegna della bibliografia, si veda Jouvet 1972). Come abbiamo riferito nel 1957 (Paasonen, MacLean e Giarman 1957), le parti cellulari dell'ippocampo, come anche i nuclei a esso strettamente associati, l'amigdala e il setto, contengono quantità relativamente alte di serotonina, e lo studio di Fuxe (1965a) ha dimostrato la presenza di terminazioni adrenergiche nello strato radiato. Riassumendo, gli studi che abbiamo passato in rassegna indicano l'esistenza di meccanismi mediante i quali l'informazione dei sistemi enterocettivo ed esterocettivo possono interagire nella formazione dell'ippocampo e influenzare l'ipotalamo e altre strutture del tronco cerebrale che svolgono una funzione nel comportamento emotivo. Il complesso striato Rimane da esaminare il problema della funzione che svolge il complesso striato (nucleo caudato più nucleo lentiforme) nel comportamento emotivo, per quanto riguarda i suoi aspetti sensoriale e percettivo. Qui siamo costretti a portare la nostra indagine in abissi di ignoranza ancor più profondi che nel caso della formazione limbica e del neocervello. Negli ultimi anni si sono accumulati numerosi dati che dimostrano come la maggior parte della corteccia limbica e della neocorteccia proietti al corpo striato (nucleo caudato più putamen, la parte laterale del nucleo lentiforme). La degenerazione osservata, tuttavia, è risultata abbastanza scarsa. Da un punto di vista evolutivo, il fatto che il lobo limbico si conformi rispetto al corpo striato un po' come uno stampo potrebbe apparire particolarmente significativo. Il rapporto apparentemente obbligato che esiste fra le due strutture è riflesso dal modo in cui la testa del nucleo caudato si prolunga in una lunga coda che è avviluppata dalla parte temporale del lobo limbico. Oltre alle connessioni reciproche con la sostanza nera, il corpo striato proietta al globo pallido (la parte mediale del nucleo lentiforme), che a sua volta stabilisce connessioni con la parte ventrale del talamo e con altre strutture del tronco cerebrale. Nauta e Mehler (1966) non sono riusciti a dimostrare l'esistenza di connessioni fra il cosiddetto tratto pallido-ipotalamico e il nucleo ventromediale o altre parti dell'ipotalamo. Sembra tuttavia che la porzione peri- pallidale della substantia innominata proietti alla regione caudo-laterale dell'ipotalamo. Albe-Fessard, Rocha-Miran- da e Oswaldo-Cruz (1960) hanno riferito che, nel gatto, afferenze di diverso tipo convergono su singoli neuroni del nucleo caudato. Non ci sono tuttavia indicazioni precise sul percorso seguito dalle vie ascendenti. Molti autori hanno notato che, nonostante centocinquant'anni di sperimentazione, si è imparato molto poco sulle funzioni del complesso striato. Il dato sperimentale secondo il quale vaste lesioni bilaterali del corpo striato o del globo pallido nei mammiferi possono non avere alcun effetto visibile sulla motilità è una prova che contraddice l'opinione clinica tradizionale secondo la quale queste strutture hanno soltanto funzioni motorie. Forse perché i ricercatori si interessano principalmente a problemi quali l'apprendimento, la memoria, la percezione e altri analoghi, non si è mai impostata la ricerca nel modo giusto. Nel nostro laboratorio di esperimenti sul campo abbiamo iniziato uno studio della funzione svolta dal complesso striato nelle forme naturali di comportamento, sottoponendo a verifica l'ipotesi che esso sia fondamentale per le forme di comportamento specifiche della specie e determinate geneticamente, come la scelta del luogo in cui stabilire la propria dimora, la presa di possesso del territorio e la sua difesa, la caccia, il ritorno a casa, l'accoppiamento, la formazione delle gerarchie sociali, la scelta dei capi. Noi abbiamo anche la speranza che questo lavoro possa servire a chiarire i meccanismi neurali che sono alla base delle varie forme di comportamento, come quello coatto, ripetitivo, ritualistico, ingannevole, imitativo. L'approccio comparato a questo studio è reso più facile e opportuno ora che gli sviluppi della neurochimica (in particolare la colorazione di Koelle per la colinesterasi e la tecnica di istofluorescenza di Falck e Hillarp) hanno reso possibile identificare le strutture corrispondenti nel complesso striato di rettili, uccelli e mammiferi. In un lavoro in cui ho studiato l'effetto di lesioni fatte in varie parti del cervello in più di novanta scimmie scoiattolo (1972a, 1973a), ho raccolto prove del fatto che il complesso striato è essenziale per l'espressione del comportamento di parata tipico della specie, come anche per i fattori imitativi associati. E’ proprio dei meccanismi imitativi il fatto che rappresentazioni parziali abbiano la capacità di scatenare forme di comportamento replicativo. Nel caso delle scimmie scoiattolo, l'immagine riflessa di un solo occhio può bastare a scatenare la parata nella sua forma completa (MacLean 1964b). Nei rettili e nelle forme inferiori, zimbelli, o anche parte di zimbelli, sono sufficienti a innescare il corteggiamento o una parata aggressiva, o altri schemi di comportamento. Nel caso dei mammiferi domestici, vengono usati zimbelli parziali per facilitare la raccolta di sperma necessario per l'inseminazione artificiale. E’ ben noto che forme indistinte o rappresentazioni parziali hanno la capacità di evocare reazioni spaventate e paranoidi negli animali e nell'uomo. Come ho ricordato all'inizio di questo capitolo, i neuropsicologi sembrano essere interessati alle illusioni percettive solo nella misura in cui queste aiutano ad analizzare i meccanismi mediante i quali i soggetti ottengono immagini perfette di oggetti indipendentemente dalle dimensioni. E’ evidente che nel lavoro neuroetologico futuro si dovrà rivolgere maggiore attenzione ai meccanismi neurali che rendono conto delle risposte comportamentali complesse alle immagini illusorie e alle rappresentazioni parziali. Come si potrebbe illustrare con numerosi esempi, il problema sussiste non solo per quanto riguarda il sistema visivo, ma anche per altri sistemi sensoriali. Riassunto Nel corso dell'evoluzione, il proencefalo dei primati si è sviluppato e si è ingrandito seguendo le linee di tre strutture di base che possiamo indicare come rettiliana, di mammifero primitivo e di mammifero recente. Queste tre formazioni evolutive, pur essendo radicalmente differenti nelle loro proprietà strutturali e chimiche, sono unite, per così dire, in un cervello trino. Questo capitolo fa il punto sulla questione del modo in cui i meccanismi sensoriale e percettivo esercitano la loro influenza sulle strutture proencefaliche che si ritengono coinvolte nell'esperienza e nell'espressione delle emozioni. È stato dato particolare rilievo ai recenti risultati sperimentali ottenuti dallo studio delle afferenze al sistema limbico, che rappresenta un'eredità che ci proviene dai mammiferi primitivi e di cui è stata dimostrata l'importante funzione nel comportamento emotivo.
|