Recentemente, nuove e promettenti ricerche ci hanno permesso di utilizzare i fossili per penetrare aspetti della biologia dei nostri progenitori estinti come, fino a qualche anno fa, non ci saremmo nemmeno immaginati. Per fare qualche esempio, ora è possibile valutare con una certa affidabilità a quale età i piccoli di una particolare specie umana venivano svezzati, a quale età raggiungevano la maturità sessuale, quale fosse l'aspettativa di vita degli individui della specie e così via. Equipaggiati dei mezzi necessari per l'acquisizione di questo genere di dati, siamo giunti alla constatazione che Homo fu un ominide che sin dall'inizio si differenziò dagli altri.
La scoperta della discontinuità biologica fra Australopithecus e Homo ha cambiato sostanzialmente la nostra interpretazione dell'evoluzione umana.
Fino alla comparsa del genere Homo, tutte le antropomorfe bipedi erano caratterizzate da encefalo piccolo, molari grandi, prognatismo e strategie di sussistenza simili a quelle delle antropomorfe ancestrali. Esse si nutrivano prevalentemente di cibo vegetale, ed è probabile che la loro organizzazione sociale rassomigliasse a quella del babbuino della savana attuale. L'unico carattere umano posseduto da queste specie - le australopitecine - era l'andatura bipede. In un'epoca precedente 2,5 milioni di anni or sono - non siamo ancora in grado di essere più precisi - si evolse la prima specie umana dotata di un cervello fortemente espanso. Anche i suoi denti erano diversi, ed è probabile che ciò riflettesse un adattamento correlato al passaggio da una dieta completamente vegetale a una che comprendeva la consumazione di carne.
Questi due aspetti dei più antichi rappresentanti del genere Homo - il cambiamento delle dimensioni encefaliche e dell'architettura dentaria - sono noti dal ritrovamento dei primi fossili di Homo habilis, avvenuto circa trentanni fa. Forse perché l'uomo moderno attribuisce un'importanza talora eccessiva alle capacità mentali, i paleoantropologi hanno organizzato gran parte della ricerca intorno al sorprendente accrescimento encefalico - da circa 450 cm3 a più di 600 cm3 - avvenuto con l'evoluzione di Homo habilis. Questo rappresentò senza dubbio una componente importante dell'adattamento che fece imboccare una nuova via all'evoluzione umana, ma ne fu soltanto un aspetto. Le nuove ricerche condotte sulle caratteristiche biologiche dei nostri progenitori hanno rivelato numerosi altri cambiamenti che contribuirono al passaggio dalla condizione «scimmiesca» a quella «umana».
Uno degli aspetti più importanti dello sviluppo di un individuo della nostra specie è che i piccoli nascono praticamente inetti e la loro infanzia dura a lungo. Inoltre, come chiunque abbia avuto figli avrà constatato, durante l'adolescenza il ragazzo attraversa un periodo di crescita improvvisa durante la quale, centimetro dopo centimetro, la sua altezza aumenta con sorprendente rapidità. In questo l'uomo non ha confronti nel mondo animale: la maggioranza dei mammiferi - scimmie antropomorfe comprese - passa quasi direttamente dall'infanzia alla condizione di adulto. Nel corso della sua improvvisa crescita l'adolescente umano - maschio o femmina che sia - acquisterà intorno al 25 per cento in statura, mentre la crescita regolare dello scimpanzé gli farà acquistare circa il 14 per cento di altezza al raggiungimento della maturità.
Barry Bogin, biologo della University of Michigan, ha elaborato una nuova interpretazione dei diversi ritmi di crescita. Il ritmo di crescita corporea del bambino è basso in confronto a quello delle scimmie antropomorfe, benché il tasso di accrescimento encefalico sia quasi uguale. Possiamo quindi affermare che i bambini sono più piccoli di quanto lo sarebbero se il loro ritmo di crescita fosse quello normalmente osservato fra le grandi scimmie. Il vantaggio evolutivo, ipotizza Bogin, ha a che fare con l'enorme quantità di nozioni che un giovane essere umano deve imparare per integrarsi nella propria cultura. Un ragazzo in crescita apprende di più dagli adulti quando vi sia una significativa differenza di mole corporea, poiché in questo caso è più facile stabilire un rapporto fra insegnante e studente. Al contrario, se i ragazzi avessero la mole corporea comportata dal ritmo di crescita proprio delle scimmie si potrebbero sviluppare rivalità. Quando il periodo dell'apprendimento culturale si è concluso, il corpo «recupera» mediante l'esplosiva crescita adolescenziale di cui abbiamo parlato.
L'uomo diviene tale, abbiamo detto, interiorizzando a ritmo serrato non solo quanto gli occorrerà sapere per sopravvivere ma anche le consuetudini sociali, i rapporti di parentela e le norme del vivere comune: in sintesi, la cultura cui appartiene. L'ambiente sociale in cui i piccoli vengono allevati e i bambini più grandi vengono educati è molto più caratteristico nell'uomo che nelle scimmie antropomorfe. Possiamo affermare che la cultura è l'adattamento umano per eccellenza, reso possibile dall'unicità del nostro modello di infanzia e di maturazione.
La condizione di totale dipendenza del neonato umano non è tanto un adattamento culturale quanto una necessità biologica. I piccoli dell'uomo vengono al mondo troppo presto, una conseguenza sia delle grandi dimensioni del nostro encefalo sia delle costrizioni comportate dall'architettura della pelvi umana. Da non molto abbiamo iniziato a renderci conto che le dimensioni del cervello influenzano ben più della mera intelligenza, e che a esse si correlano i fattori di sviluppo degli individui di una specie, come l'età di svezzamento, l'età in cui raggiungeranno la maturità sessuale, la lunghezza del periodo di gestazione e la longevità. Nelle specie caratterizzate da un cervello più grande, questi fattori tendono a impegnare periodi più lunghi: i piccoli vengono svezzati più tardi e anche la maturità sessuale viene raggiunta più tardi, la gestazione è più lunga e gli individui vivono più a lungo. Un semplice calcolo basato sul confronto con altri primati rivela che la lunghezza della gestazione in Homo sapiens, il cui volume endocranico medio è di 1350 cm3, dovrebbe essere di ventun mesi, e non di nove quale è di fatto. Ne consegue che, quando viene al mondo, il neonato umano ha un anno di crescita da recuperare, da cui la sua totale inettitudine.
Ma perché questo adattamento? Perché la natura ha esposto i nostri piccoli ai pericoli insiti in una nascita troppo precoce? La risposta ci proviene da alcune considerazióni sulle dimensioni encefaliche dell'uomo e delle antropomorfe. Il volume endocranico medio di una scimmia antropomorfa appena nata è di circa 200 cm3, la metà del valore che essa raggiungerà da adulta. Il raddoppiamento avverrà rapidamente e a uno stadio precoce della sua vita. L'encefalo di un neonato umano, al contrario, ha un volume di circa un terzo di ciò che sarà da adulto, e si triplicherà con una crescita rapida e precoce. L'uomo e le antropomorfe, dunque, sono accomunati dalla precocità del loro accrescimento encefalico; ne consegue che se il volume encefalico dei neonati umani dovesse raddoppiarsi - come avviene fra le antropomorfe - il valore alla nascita dovrebbe aggirarsi intorno ai 675 cm3. Ma, come molte donne sanno, è già abbastanza difficoltoso, e talvolta pericoloso, dare alla luce un bambino con un volume encefalico normale, che si aggira intorno a 385 cm3. In realtà, l'apertura della pelvi si è ingrandita nel corso dell'evoluzione umana proprio per far fronte all'accrescimento encefalico. Ma vi era un limite meccanico a questo cambiamento - un limite imposto dalla necessità di un'efficiente locomozione bipede - che quando il volume encefalico medio del neonato umano giunse al valore attuale non potè più essere superato.
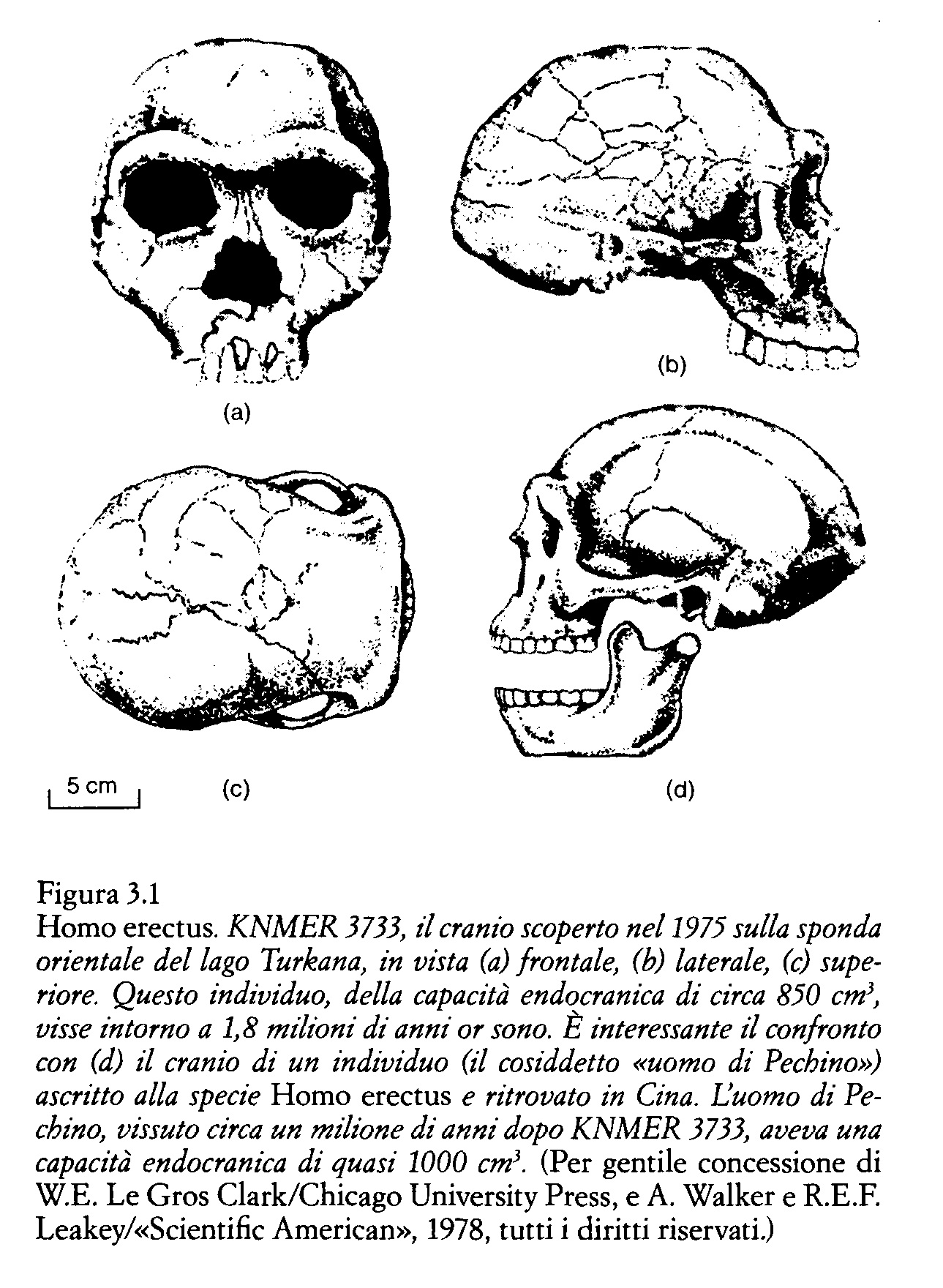
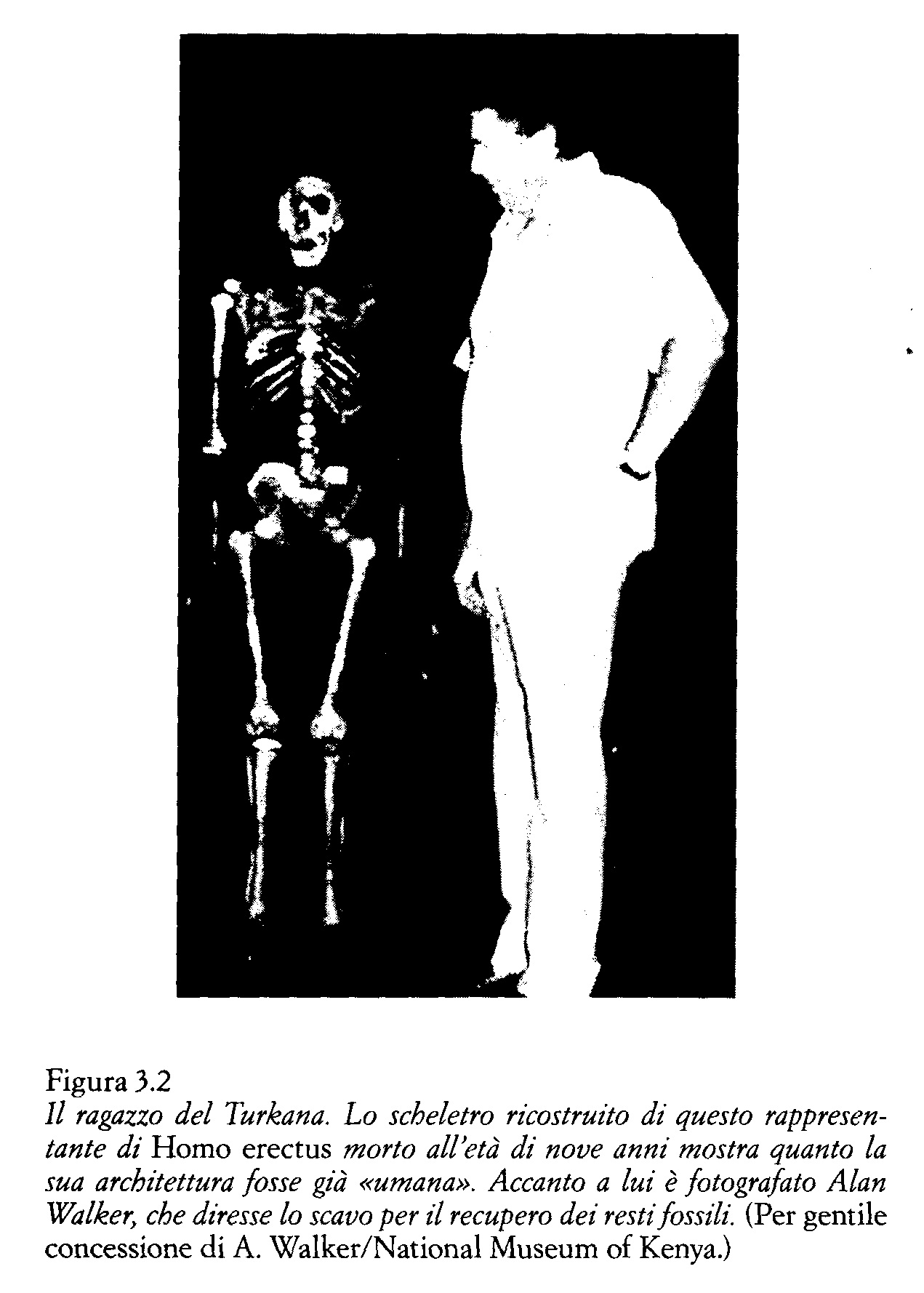
Dal punto di vista evolutivo possiamo affermare che, in linea di massima, l'uomo iniziò a discostarsi dal modello di accrescimento delle antropomorfe quando il volume encefalico degli individui adulti superò i 770 cm3. Oltre questo valore, il volume endocranico si sarebbe più che raddoppiato rispetto al valore alla nascita, dando inizio al modello di «inettitudine» del neonato umano che viene alla luce «troppo presto». Homo habilis, il cui volume encefalico da adulto è di circa 800 cm3, sembra collocarsi al limite fra il modello che caratterizza le antropomorfe e quello umano, mentre il volume dei più antichi resti di Homo erectus, che si aggira intorno a 900 cm3, segna l'avvio di questa specie verso il valore medio dell'uomo attuale. Ma non dobbiamo dimenticare che il nostro è un ragionamento «in linea di principio», condotto presumendo che nelle femmine di Homo erectus il canale del parto avesse le stesse dimensioni che nella donna attuale. In pratica abbiamo compreso più chiaramente quanto, sotto questo aspetto, Homo erectus fosse già simile all'uomo attuale studiando le misure della pelvi del ragazzo del Turkana, lo scheletro ritrovato dalla mia équipe verso la metà degli anni Ottanta sulla riva occidentale del lago da cui ha preso il nome.
Nell'uomo attuale, l'apertura della pelvi è più o meno uguale nei maschi e nelle femmine. Conseguentemente, prendendone le misure sullo scheletro del ragazzo del Turkana abbiamo ottenuto un valore abbastanza preciso del canale del parto di sua madre. Il mio amico e collega Alan Walker, anatomista della Johns Hopkins University, ha ricostruito la pelvi del ragazzo a partire dai resti scheletrici ritrovati. Dalla successiva misurazione è scaturito che l'apertura pelvica era inferiore a quella di Homo sapiens, e che il volume endocranico dei neonati di Homo erectus poteva aggirarsi intorno a 275 cm3, un valore considerevolmente inferiore a quello dei neonati umani attuali.cm3
Le implicazioni di questa scoperta sono evidenti. I neonati di Homo erectus nascevano con un volume encefalico pari a un terzo del valore da adulti, come avviene nell'uomo attuale, e, come avviene nell'uomo attuale, dovevano necessariamente venire al mondo del tutto inetti. Possiamo quindi dedurre che le assidue cure parentali che fanno parte della cultura umana attuale avevano già iniziato a svilupparsi con la comparsa di Homo sapiens, circa 1,7 milioni di anni or sono.
Riguardo a Homo habilis - il progenitore immediato di Homo erectus - non possiamo fare gli stessi calcoli perché non abbiamo mai ritrovato ossa pelviche. Tuttavia è lecito ipotizzare che se i piccoli di Homo habilis nascevano con un volume encefalico pari a quello dei piccoli di Homo erectus, allora anch'essi avrebbero avuto bisogno di nascere «troppo presto», anche se non altrettanto presto; inoltre, anch'essi sarebbero stati inetti alla nascita, quantunque non così a lungo, e infine anch'essi avrebbero avuto bisogno di assidue cure parentali, ma in misura inferiore. Sembra dunque che il genere Homo avesse imboccato la direzione dell'uomo attuale fin dall'inizio. Lo stesso tipo di ragionamento può essere applicato alle specie australopitecine, che, avendo un volume encefalico confrontabile con quello delle scimmie antropomorfe, avranno presumibilmente seguito il loro stesso modello di sviluppo precoce.
Un lungo periodo infantile durante il quale erano indispensabili assidue cure parentali era già caratteristico delle più antiche forme del genere Homo: questo, perlomeno, è quanto abbiamo concluso. Ma che dire del resto dell'infanzia? In quale epoca iniziò a prolungarsi per permettere al bambino di impadronirsi di nozioni sia pratiche che culturali, cui sarebbe seguita un'adolescenza caratterizzata da quella repentina crescita di cui abbiamo già parlato.
Nell'uomo attuale il prolungamento dell'infanzia viene conseguito mediante la riduzione del ritmo di crescita rispetto a quello delle scimmie antropomorfe, e quindi il bambino raggiunge alcuni momenti significativi del suo sviluppo - come ad esempio l'eruzione dei denti - più tardi di quanto non facciano i piccoli delle grandi scimmie. Il primo molare permanente compare a sei anni circa nel bambino e a tre anni nei piccoli delle antropomorfe, il secondo molare compare rispettivamente fra undici e dodici anni e a sette anni, e il terzo rispettivamente fra diciotto e vent'anni e a nove anni. Per rispondere all'interrogativo sul quando l'infanzia umana iniziò a prolungarsi avevamo bisogno di disporre di mascellari fossili per determinare l'età di eruzione dei molari.
Il ragazzo del Turkana, ad esempio, morì appena dopo l'eruzione del secondo molare. Se Homo erectus avesse seguito il modello di lento sviluppo infantile tipico dell'uomo attuale, alla morte avrebbe avuto circa undici anni. Al contrario, se la sua specie avesse seguito uno schema di crescita simile a quello delle antropomorfe, ne avrebbe avuti sette.
All'inizio degli anni Settanta, Alan Mann, della University of Pennsylvania, compì un'estesa analisi di denti fossili umani e concluse che tutte le specie di Australopithecus e di Homo seguivano il modello di lenta crescita durante l'infanzia osservato nell'uomo attuale. Il suo lavoro esercitò una grande influenza e suffragò il già diffuso luogo comune secondo cui tutte le specie ominidi, australopitecine comprese, seguivano il modello umano attuale. E in effetti quando trovammo la mandibola del ragazzo del Turkana e constatai che l'eruzione del secondo molare era già avvenuta pensai subito che fosse morto a undici anni, l'età che avrebbe avuto se fosse appartenuto alla specie Homo sapiens. D'altro canto, anche al bambino di Taung, un individuo della specie Australopithecus africanus, era stata inizialmente attribuita un'età di sette anni perché il primo molare era in fase di eruzione.
Ma sul finire degli anni Ottanta il lavoro di diversi ricercatori fece vacillare queste convinzioni. Holly Smith, un'antropologa della University of Michigan, mise a punto una tecnica per dedurre il modello di sviluppo degli individui rappresentati dai fossili umani, che si avvaleva della correlazione fra volume encefalico ed età di eruzione del primo molare. Per prima cosa, Smith raccolse quanti più dati potè sia sull'uomo che sulle antropomorfe, dopo di ehe studiò una serie di fossili umani per stabilire quali correlazioni intercorressero fra loro. Ne emersero tre modelli di sviluppo: quello dell'uomo attuale, nel quale l'eruzione del primo molare avviene a sei anni e la cui aspettativa di vita è di sessantasei anni circa, quello delle antropomorfe, con valori di poco più di tre anni e quarantanni rispettivamente, e in ultimo un modello intermedio. I rappresentanti di Homo erectus successivi al ragazzo del Turkana, cioè gli individui vissuti intorno a 800.000 anni or sono, seguivano il modello umano attuale, come anche i neandertaliani. Tutte le australopitecine, invece, rientravano nel modello delle scimmie antropomorfe. I più antichi rappresentanti di Homo erectus - come il ragazzo del Turkana - seguivano il modello intermedio: era probabile che il primo molare fosse spuntato quando il ragazzo aveva poco più di quattro anni e mezzo, e se egli non avesse incontrato una morte prematura avrebbe avuto un'aspettativa di vita di circa cinquantadue anni.
I modelli di Smith dimostrarono che il ritmo di crescita delle australopitecine non è simile a quello dell'uomo moderno ma è paragonabile a quello delle scimmie antropomorfe. Ella ha altresì dimostrato che i più antichi rappresentanti di Homo erectus seguivano un modello di crescita intermedio fra quello dell'uomo attuale e quello delle antropomorfe, e ora noi possiamo concludere che il ragazzo del Turkana, quando morì, aveva nove anni e non undici come si era supposto inizialmente.
Poiché queste conclusioni erano contrarie a quanto sostenuto da una generazione di paleoantropologi, scoppiò un'accesa controversia. Naturalmente c'era la possibilità che Smith avesse commesso un errore. In queste circostanze un altro studio che rappresenti una sorta di prova del nove è sempre ben accetto, e infatti arrivò prontamente. Gli anatomisti Christopher Dean e Tim Bromage, che all'epoca lavoravano presso lo University College di Londra, studiarono un metodo di valutazione diretta dell'età di eruzione dei denti basato sul principio che nello stesso modo in cui è possibile contare gli anelli di accrescimento nel tronco di un albero è possibile contare le microscopiche linee di crescita di un dente. In pratica questo metodo di calcolo non è così semplice, anche perché talvolta sorgono incertezze sul modo in cui le linee si sono formate. Ciononostante, Dean e Bromage provarono ad applicare la loro tecnica alla mandibola di un'australopitecina identica a quella del bambino di Taung in termini di sviluppo dentario, scoprendo che l'individuo era morto a poco più di tre anni, proprio quando il suo primo molare era in fase di eruzione, e che quindi si collocava entro il modello di crescita delle antropomorfe.
Quando Dean e Bromage studiarono una serie di altri denti fossili umani individuarono, come già aveva fatto la Smith, tre modelli: quello dell'uomo attuale, quello delle antropomorfe e un modello intermedio. Ancora una volta, le australopitecine si situavano nel modello delle antropomorfe, Homo erectus tardivo e i neandertaliani si situavano nel modello dell'uomo attuale e i rappresentanti più antichi di Homo erectus seguivano il modello intermedio. E ancora una volta i risultati suscitarono polemiche, soprattutto intorno alla questione delle australopitecine, che alcuni avrebbero preferito situare non entro il modello di crescita delle antropomorfe ma in quello dell'uomo attuale.
Il dibattito venne concluso quando l'antropologo Glenn Conroy e il clinico Michael Vannier, entrambi della Washington University, St. Louis, applicarono a questo campo di ricerca un'avanzata tecnologia diagnostica. Utilizzando la tomografia computerizzata (Tac) essi poterono studiare l'interno della mandibola fossile del bambino di Taung e sostanzialmente confermarono le conclusioni di Dean e Bromage: egli era morto a poco meno di tre anni, e il suo modello di crescita si era rivelato quello delle scimmie antropomorfe.
La possibilità di inferire dati di natura biologica da resti fossili mediante l'analisi degli schemi di sviluppo sia corporeo che dentario è di grande importanza per la paleoantropologia, poiché permette a questa disciplina di «rimpolpare» le informazioni offerte dalle ossa fossili. Per esempio, possiamo affermare che il ragazzo del Turkana venne probabilmente svezzato poco prima che compisse quattro anni, e che, se fosse vissuto sarebbe diventato sessualmente maturo a circa quattordici anni. E probabile che sua madre avesse avuto il primo figlio a circa tredici anni, e che la gravidanza fosse durata nove mesi; in seguito sarebbe rimasta gravida ogni tre o quattro anni. Questi modelli ci indicano che al tempo dei più antichi rappresentanti di Homo erectus certe caratteristiche biologiche dei progenitori dell'uomo si stavano già evolvendo verso quelle dell'uomo attuale e, soprattutto, stavano divergendo dal modello proprio delle scimmie antropomorfe, mentre le australopitecine continuavano ad aderire al modello di queste ultime.
Il mutamento evolutivo compiuto dai più antichi rappresentanti del genere Homo in direzione dei modelli di crescita e di sviluppo dell'uomo attuale si compì nell'ambito di un contesto sociale. Tutti i primati sono animali sociali, ma l'uomo attuale ha sviluppato questa caratteristica al massimo grado. I mutamenti biologici inferiti mediante lo studio dei denti delle più antiche forme di Homo ci dicono che le interazioni sociali fra gli individui di questa specie avevano già incominciato a intensificarsi, creando un ambiente favorevole alla germinazione della cultura. A quanto sembra, anche l'organizzazione sociale subì cambiamenti di rilievo. Ma come possiamo esserne certi? La certezza ci deriva dal confronto della mole corporea di maschi e femmine e da quanto abbiamo appreso circa queste differenze studiando specie di primati attuali, come babbuini e scimpanzé.
Come abbiamo già detto, fra i babbuini della savana la taglia dei maschi è doppia rispetto a quella delle femmine. Ora i primatologi sanno che una differenza di taglia così accentuata ricorre quanto più violenta è la competitività fra i maschi sessualmente maturi per l'accoppiamento. Come nella maggior parte delle specie di primati, il maschio di babbuino che ha raggiunto la maturità lascia il gruppo natale per unirsi a un altro, che spesso occupa un territorio contiguo, dopo di che si troverà per sempre in competizione con i maschi già presenti nel gruppo. Questo modello di migrazione maschile fa sì che nella maggior parte dei gruppi i maschi non siano legati da rapporti di parentela, motivo per cui non hanno alcuna ragione «darwiniana» (cioè genetica) di cooperare fra loro.
Fra gli scimpanzé, invece, per ragioni non ancora ben comprese i maschi rimangono nel loro gruppo natale e le femmine emigrano. Ne consegue che gli scimpanzé maschi hanno valide ragioni darwiniane per cooperare nell'acquisizione delle femmine, perché, in quanto fratelli, hanno la metà del patrimonio genetico in comune. Essi collaborano per difendersi da altri gruppi di conspecifici e in occasionali spedizioni di caccia, nel corso delle quali solitamente cercano di isolare e uccidere una scimmia inferiore di piccola taglia. La relativa assenza di competitività e il maggior spirito di collaborazione osservati fra gli scimpanzé sono riflessi dal fatto che la taglia dei maschi è appena del 15-20 per cento superiore a quella delle femmine.
Quanto alla mole corporea, i maschi delle australopitecine si collocano entro il modello dei babbuini ed è quindi ipotizzabile che la loro vita sociale fosse organizzata in modo analogo a quanto osserviamo fra i gruppi di babbuini attuali. Quando il ritrovamento di resti fossili ci consente di confrontare la mole corporea di maschi e femmine rappresentanti delle più antiche forme di Homo, salta immediatamente all'occhio che si era già verificato un significativo cambiamento: la taglia dei maschi superava quella delle femmine per non più del 20 per cento, come fra gli scimpanzé. Come hanno sostenuto Robert Foley e Phyllis Lee, antropologi della Cambridge University, il cambiamento nella differenza di mole corporea verificatosi all'origine del genere Homo rispecchia certamente un cambiamento intervenuto nell'organizzazione sociale. E' molto probabile che fra i primi rappresentanti di Homo i maschi rimanessero nel gruppo natale con i propri fratelli e fratellastri e che le femmine si trasferissero in altri gruppi. Le relazioni di parentela, come ho già detto, favoriscono lo spirito di cooperazione fra i maschi.
Non possiamo indicare con certezza che cosa diede l'avvio ai mutamenti avvenuti nell'organizzazione sociale, ma, sicuramente, una maggiore cooperazione fra i maschi doveva comportare notevoli vantaggi. Alcuni paleoantropologi sostengono che in questo periodo la necessità di difendersi dai gruppi che occupavano territori confinanti si era fatta pressante. E' fortemente probabile che si fossero verificati anche alcuni cambiamenti nelle necessità economiche. Numerose testimonianze concorrono a indicare che intervenne un mutamento nell'alimentazione degli individui del genere Homo, e che la carne stava divenendo una fonte energetica sempre più importante. I cambiamenti osservati nell'architettura dentaria dei più antichi rappresentanti di Homo indicano che essi si nutrivano di carne, una conclusione confermata dalla complessità raggiunta nello stesso periodo dalla tecnologia litica. Inoltre non possiamo escludere che il considerevole accrescimento encefalico documentato dai resti fossili avesse imposto alla specie di integrare la propria alimentazione con una ricca fonte energetica quale è la carne.
Come ogni biologo sa bene, il funzionamento del cervello comporta alti costi metabolici in termini di energia. Nell'uomo attuale, per esempio, il peso del cervello ammonta appena al 2 per cento del peso corporeo ma consuma il 20 per cento dell'energia totale. Fra tutti i mammiferi i primati costituiscono il gruppo che ha il cervello più grande, una caratteristica enormemente accentuatasi nell'uomo, il cui cervello pesa tre volte quello di un'antropomorfa di uguale taglia. Robert Martin dell'Istituto di Antropologia di Zurigo ha fatto osservare che l'accrescimento della massa cerebrale potè verificarsi solo in seguito a un aumento dell'apporto energetico, e che l'alimentazione degli individui del genere Homo avrebbe dovuto essere non solo regolare ma nutrizionalmente ricca, e la carne è appunto una ricca fonte di calorie, proteine e grassi. I più antichi membri del genere umano avrebbero potuto «permettersi» l'accrescimento cerebrale che li avrebbe portati a superare i valori osservati nelle australopitecine solo aggiungendo alla propria dieta una buona proporzione di proteine animali.
Per tutte le ragioni che ho esposto, ritengo che il più importante adattamento evolutivo verificatosi agli inizi del genere Homo sia stata la consumazione regolare di una certa quantità di carne. Se i nostri progenitori catturassero prede vive o si limitassero a sfruttare le carcasse abbandonate in cui si imbattevano - o se attuassero entrambe le strategie - è una questione vivacemente dibattuta fra i paleoantropologi, come vedremo nel prossimo capitolo. Ma io non dubito dell'importanza che la carne ebbe nella loro vita quotidiana. Inoltre è probabile che il passaggio dalla semplice ricerca di cibo vegetale all'approvvigionamento di carne abbia imposto ai primi gruppi umani la necessità di sviluppare un certo grado di organizzazione sociale e di cooperazione.
Sappiamo bene che quando nella strategia di sussistenza di una specie si verifica un cambiamento di rilievo, solitamente a questo ne seguono altri. Molto spesso i cambiamenti secondari riguardano la sua anatomia, e intervengono nel corso dell'adattamento alla nuova alimentazione. Come abbiamo visto, l'architettura dentaria e lo scheletro facciale dei più antichi rappresentanti del genere Homo sono diversi da quelli delle australopitecine, probabilmente come adattamento a una dieta carnea.
Una delle più recenti conclusioni cui i paleoantropologi sono giunti è che, oltre alle differenze osservate nella dentatura, i primi rappresentanti del genere Homo erano fisicamente molto più attivi delle australopitecine. Due ricerche condotte indipendentemente indicano che questi nostri progenitori erano corridori molto efficienti, e che furono la prima specie umana ad acquisire questa capacità.
Alcuni anni fa l'antropologo Peter Schmid, collega di Robert Martin presso l'Università di Zurigo, ebbe l'occasione di poter studiare lo scheletro della famosa Lucy. Utilizzando calchi in fiberglass, Schmid iniziò a ricomporre lo scheletro in base al presupposto che la sua anatomia fosse pienamente umana. Invece rimase sorpreso da alcune constatazioni: la gabbia toracica si rivelò di forma conica, come nelle antropomorfe, e non di forma più o meno cilindrica come nell'uomo, e c'era da ritenere che anche la presumibile morfologia esterna della spalla e del tronco e l'assenza di restringimento in corrispondenza della vita le conferissero un aspetto spiccatamente scimmiesco.
Nel corso di un incontro internazionale svoltosi a Parigi nel 1989, Schmid descrisse le importanti implicazioni della propria scoperta. «Australopithecus afarensis» affermò «non era in grado di espandere il torace per respirare così profondamente come facciamo noi durante la corsa. Il suo addome sporgente e l'assenza di restringimento alla vita limitavano quella flessibilità che invece è essenziale per correre come l'uomo.» Se ne deduce che i rappresentanti del genere Homo erano corridori, ma non quelli del genere Australopithecus.
Una seconda testimonianza sulla questione dell'agilità delle australopitecine è scaturita dallo studio sul rapporto fra peso corporeo e statura condotto da Leslie Aiello, che compì misurazioni sia nell'uomo che nelle antropomorfe attuali e le confrontò con dati tratti dallo studio di fossili umani. Le antropomorfe attuali sono di costituzione molto robusta rispetto alla loro statura, e pesano il doppio di un uomo della stessa altezza. Anche lo studio dei fossili fornì dati che riproponevano un modello ormai noto: la costituzione delle australopitecine era antropomorfa mentre quella di tutte le specie ascritte a Homo seguiva il modello umano. Sia i dati di Leslie Aiello che lo studio anatomico di Peter Schmid sono congruenti con la differenza - osservata da Fred Spoor - fra l'architettura dell'orecchio interno delle australopitecine e quella di individui ascritti a Homo: una maggiore attitudine al bipedismo deve accompagnarsi a un innalzamento della statura.
Durante l'evoluzione umana, come abbiamo già detto, oltre all'accrescimento cerebrale si verificarono altri cambiamenti. Ora possiamo analizzarli meglio: le australopitecine erano bipedi ma la loro agilità era limitata, mentre i rappresentanti delle specie di Homo erano atletici.
Il bipedismo, dunque, iniziò a evolversi come un più efficiente modo di locomozione in un ambiente fisico mutato, e consentì a un'antropomorfa bipede di sopravvivere in un habitat inadatto a quelle che invece avevano mantenuto un genere di vita più convenzionale. Le antropomorfe bipedi, infatti, durante la ricerca del cibo avevano la possibilità di percorrere spazi aperti molto più estesi. Con l'evoluzione del genere Homo si sviluppò una nuova forma di locomozione bipede che presupponeva una maggiore agilità e una più intensa attività fisica. L'agilità e la statura dell'uomo attuale gli permettono una deambulazione sicura ed efficiente e un'adeguata dispersione del calore corporeo, fattori entrambi importanti per animali che, come i più antichi rappresentanti del genere Homo, conducevano una vita attiva in ambienti caldi e aperti. Un'efficiente e sicura camminata bipede rappresentò un adattamento cruciale per l'evoluzione degli ominidi. In questo cambiamento, come vedremo nel prossimo capitolo, ebbe una grande importanza la caccia, che si ritiene venisse praticata, almeno in qualche misura.
In un animale attivo, la capacità di disperdere il calore è importante per la fisiologia del cervello, una questione più volte trattata dall'antropologa Dean Falk della State University of New York, Albany. In una ricerca svolta negli anni Ottanta sul cervello dei rappresentanti del genere Homo ella dimostrò che il sistema di drenaggio dei vasi sanguigni consentiva un efficiente raffreddamento, mentre nelle australopitecine ciò avveniva in misura inferiore. La cosiddetta «ipotesi del radiatore» formulata dalla Falk è un'ulteriore dimostrazione di quanto fosse ampio l'adattamento del genere Homo.
È superfluo sottolineare che l'adattamento del genere Homo fu un successo evolutivo, testimoniato dalla nostra stessa esistenza. Ma perché insieme a noi non si è evoluta qualche altra antropomorfa bipede?
Nell'Africa meridionale e orientale di 2 milioni di anni or sono, Homo coesisteva con diverse specie di Australopithecus, ma un milione di anni dopo le australopitecine si erano estinte ed egli si trovò in una condizione di «splendido isolamento». (Noi tendiamo a considerare l'estinzione come un fallimento evolutivo, una disavventura che capita alle specie che per qualche motivo soccombono alle sfide lanciate dalla natura, e non pensiamo invece che il 99,9 per cento di tutte le specie finora esistite è attualmente estinto, probabilmente a causa sia della cattiva sorte che dei cattivi geni.) Che cosa sappiamo del destino toccato alle australopitecine?
Spesso mi domandano se penso che il genere Homo, essendo diventato carnivoro, potrebbe essersi cibato anche di questi parenti prossimi, portandoli all'estinzione. Io non dubito che occasionalmente le più antiche popolazioni umane uccidessero le australopitecine per qualche motivo vulnerabili in cui si imbattevano, nello stesso modo in cui non appena potevano uccidevano un'antilope o qualsiasi altra preda. Ma la causa della loro estinzione è probabilmente più banale.
Noi sappiamo che Homo erectus ebbe un grandioso successo evolutivo in quanto fu la prima specie umana a diffondersi fuori dell'Africa. E quindi probabile che i suoi membri si fossero moltiplicati tanto rapidamente da poter competere con le australopitecine per una risorsa essenziale alla loro sopravvivenza: il cibo. Inoltre, fra 2 milioni e un milione di anni or sono, anche le scimmie che vivevano al suolo - cioè i babbuini - ebbero un grande successo evolutivo e riproduttivo, ed è probabile che anch'esse abbiano contribuito alla competizione. Non è quindi da escludere che le australopitecine abbiano dovuto soccombere alla duplice pressione esercitata dalle popolazioni umane e da quelle di babbuini.