
In questo articolo famosissimo, pubblicato nel 1979 nei Proceedings della Royal Society, serie B, volume 205, n. 1161, c’è la contestazione critica dell’ideologia adattamentista, di cui ho tenuto gran conto nella stesura del saggio sulle emozioni (Il mostro di belle speranze). Per quanto la lettura possa risultare un po’ ostica, data la terminologia scientifica, essa è utilissima per capire il dibattito che si è aperto ed è tuttora in corso in seno all’evoluzionismo darwiniano.
Ne riassumo brevemente i termini.
Dopo avere accumulato una serie consistente e poco confutabile di prove che gli hanno consentito di trasformare l’evoluzionismo in una teoria scientifica, laddove fino allora era stata solo un’intuizione filosofica, Darwin si è trovato di fronte al problema di fornire una spiegazione del fenomeno. Esclusa una creazione divina e il riferimento ad un élan vital, vale a dire ad una spinta teleologica, entrambi incompatibili con la sua mentalità, occorreva avanzare un’ipotesi che consentisse di spiegare il mistero di un processo che dalle forme più semplici di vita è giunto all’uomo: un’ipotesi, dunque, materialistica che, all’epoca non poteva avvalersi della genetica e della paleontologia.
La selezione naturale è stata, infine, tra le tante vagliate, quella accreditata da Darwin sulla base dell’unico dato disponibile: la varietà degli individui all’interno di una specie. E’ certo che, nel formulare tale ipotesi, egli abbia tenuto conto della pratica degli incroci a livello di allevamento degli animali. Dagli incroci, però, nascono nuove razze, non nuove specie.
Nell’ottica darwiniana, la speciazione è un processo lentissimo (che può durare centinaia di migliaia o milioni di anni) che si basa sull’accumulo progressivo di minuscoli piccoli cambiamenti di strutture organiche e di funzioni che vengono selezionate in virtù del loro valore adattivo e, infine, sommandosi tra loro danno luogo alla nascita di una nuova specie riproduttivamente isolata rispetto alle preesistenti.
In virtù della sua lentezza, il processo di speciazione postula una quantità indefinita di forme intermedie destinate poi a scomparire.
Darwin non aveva dubbi riguardo al fatto che lo sviluppo della paleontologia avrebbe fornito le prove dell’esistenza di queste forme intermedie.
La nascita e lo sviluppo della genetica ha confermato scientificamente, attraverso la presa d’atto che i geni sono unità discrete e la scoperta del fenomeno delle mutazioni spontanee, la sostanziale validità della teoria darwiniana. Il neodarwinismo, che rappresenta per l’appunto l’integrazione tra evoluzionismo e genetica, ha avallato sia il principio del più adatto (rispetto ad un determinato ambiente) sia il principio del gradualismo, per cui la speciazione avviene in tempi sterminatamente lunghi.
Il problema sulla validità assoluta della teoria della selezione naturale si è posto nel momento in cui la paleontologia, nonostante il suo rapido sviluppo, non solo non ha fornito prove sufficienti dell’esistenza delle forme intermedie, ma ha posto di fronte ad indizi che attestano che l’evoluzione ha riconosciuto lunghi periodi di ristagno intervallati da periodi di speciazione addirittura esuberanti.
Nonostante queste lacune, il neodarwinismo è rimasto fermo al gradualismo darwiniano e ha tentato di aggirare il problema con spiegazioni ad hoc, scientificamente insufficienti. In questa cornice, il principio dell’adattamento si è trasformato nell’ideologia adattamentista, secondo la quale tutte le strutture e le funzioni organiche in tanto esistono in quanto esse sono state selezionate originariamente sulla base del loro valore adattivo.
Nell’articolo riportato, Gould e Lewontin attaccano frontalmente il dogma adattamentista, proponendo un’ipotesi alternativa: quella per cui, se rimane vero che gran parte delle strutture biologiche sono frutto della selezione naturale, si può pensare che alcune di esse contengano potenzialità funzionali che, all’epoca in cui sono state selezionate, non servivano a nulla o servivano ad assolvere altre funzioni, e che solo successivamente hanno assunto un significato adattivo.
Immediatamente, la straordinaria novità di questa ipotesi può rimanere oscura. Essa di fatto introduce, nella cornice di una teoria meccanicistica, un grado di “creatività” del tutto casuale, che io ritengo abbia trovato la sua massima espressione nell’uomo, con il suo strano cervello nel quale potenzialità adattive e potenzialità apparentemente disadattive (come l’ansia esistenziale o le emozioni) convivono.
Al di là di questo, l’articolo di Gould e di Lewontin è un autentico gioiello, che dà la misura della “creatività” della scienza (che, forse, è un effetto casuale di un cervello che originariamente sarebbe dovuto servire solo a sopravvivere e a farsi un’idea dell’ambiente).
Un programma adattamentista ha dominato il pensiero evoluzionistico in Inghilterra e negli Stati Uniti nel corso degli ultimi quarant'anni. Esso, basato sulla fiducia che la selezione naturale agisca da fattore ottimizzante, procede scindendo un organismo in “tratti” unitari e proponendo una storia adattiva per ciascuno di essi considerato separatamente. I compromessi fra domande selettive antagoniste rappresentano il solo freno alla perfezione. Anche l'imperfezione può essere dunque considerata come il risultato di un adattamento. Noi critichiamo un tale approccio e cerchiamo di riaffermare l’idea alternativa (a lungo popolare nell'Europa continentale) che gli organismi debbano essere analizzati come insiemi integrati, con Baupläne1 determinati dall'eredità filetica, dalle modalità di sviluppo e dall'architettura generale, così che i vincoli stessi divengano più interessanti e importanti nel delineare le linee evolutive, rispetto alla forza selettiva che può mediare il cambiamento, qualora esso occorra.
Rifiutiamo il programma adattamentista per il suo fallimento nel distinguere tra l'utilità corrente e le ragioni dell'origine (i tirannosauri maschi potrebbero aver usato le loro minuscole zampe anteriori per titillare i loro partner femminili, ma ciò non spiega perché esse fossero così piccole); per la sua riluttanza a considerare delle spiegazioni alternative alle storie adattive; per la sua fiducia nella sola plausibilità come criterio di scelta fra narrazioni meramente speculative; e infine per la mancanza di una adeguata considerazione di temi antagonisti, come la fissazione casuale di alleli, la produzione di strutture non-adattive mediante correlazioni di sviluppo legate a caratteristiche selezionate (allometria, pleiotropia, compensazione materiale, correlazione meccanica forzata), la disgiunzione di adattamento e selezione, i picchi adattivi multipli e l'utilità corrente come epifenomeno concernente strutture nonadattive. Sosteniamo dunque l'approccio pluralistico proprio di Darwin nella ricerca dei fattori del cambiamento evolutivo.
1 Bauplan/ pl. Baupläne s.m. (costituito da bau – costruzione, architettura, struttura, o anche edificio, fabbricato, costruzione; plan – progetto, piano, programma, disegno, pianta) va inteso in questo caso come "piano di costruzione" di un organismo, riferendosi con ciò al suo tipo di organizzazione morfologica, fisiologica e comportamentale. Si preferisce mantenere il termine in tedesco, nelle sue diverse ricorrenze all'interno del saggio, per conservare la scelta già fatta dagli autori rispetto all'inglese. [N.d.T.]
La grande cupola centrale della Basilica di San Marco a Venezia presenta nell'organizzazione dei suoi mosaici dettagli iconografici che esprimono tutta la maestosità della fede cristiana. Tre serie di figure disposte su anelli concentrici si susseguono a partire dall'immagine centrale del Cristo: angeli, discepoli e virtù. Ciascun anello è diviso in quadranti, sebbene la cupola abbia in sé una struttura a simmetria radiale. Ogni quadrante incontra uno dei quattro pennacchi fra gli archi al di sotto della cupola.
I pennacchi – gli spazi triangolari affusolati formati all'intersezione di due archi a tutto sesto contigui e disposti in pianta ad angolo retto (Figura 1) – sono necessari sottoprodotti architettonici dell'edificazione di una cupola su archi a tutto sesto. Ciascun pennacchio contiene illustrazioni ammirevoli e fitte nel suo spazio affusolato. In uno di essi un evangelista siede nella parte più alta, fiancheggiato dalla città celeste. In basso, un uomo rappresenta uno dei quattro fiumi biblici (Tigri, Eufrate, Indo e Nilo) e versa acqua da una brocca nello stretto spazio sotto i suoi piedi.
Il risultato è così elaborato, armonioso e denso di significati che si è tentati di vederlo come il punto di partenza di ogni possibile interpretazione, cioè come la causa in qualche senso di tutta l'architettura circostante. Ma questo capovolgerebbe la corretta modalità di analisi. Il tutto ha origine infatti da un vincolo architettonico: la necessità dei quattro pennacchi e la loro forma triangolare rastremata. Si tratta certamente di spazi utilizzati dai mosaicisti nel loro lavoro, ma ciò non toglie che siano semplicemente il risultato della struttura quadripartita al di sotto della cupola.
L'architettura è piena di vincoli strutturali, e noi li cogliamo facilmente perché non imponiamo loro i nostri pregiudizi biologici. Ogni soffitto con volta a ventaglio deve avere una serie di spazi aperti lungo le linee mediane della volta, dove i bordi dei ventagli che sovrastano i pilastri si intersecano fra di loro (Figura 2). Poiché questi spazi devono per forza esistere, essi vengono spesso utilizzati per ingegnosi effetti ornamentali.
Nella cappella del King's College di Cambridge, ad esempio, essi contengono rosoni abbelliti dalla rosa dei Tudor o da una saracinesca di castello, in maniera alternata. In un certo senso la struttura risultante rappresenta un “adattamento”, ma il vincolo architettonico è chiaramente primario. Gli spazi nascono come necessari sottoprodotti delle volte a ventaglio; il loro uso appropriato è un effetto secondario. Chiunque tentasse di affermare che la struttura esiste perché l'alternarsi di abbellimenti come una rosa e una saracinesca rende tanto bella e significativa una cappella Tudor cadrebbe nella stessa ridicolaggine che Voltaire attribuiva al Dr. Pangloss: «Le cose non possono essere diverse da come sono. [...] Ogni cosa è fatta per uno scopo. I nostri nasi sono fatti per portare gli occhiali, così abbiamo gli occhiali. Le gambe sono di certo fatte per portare i pantaloni, e noi li portiamo».
Il fatto è che gli evoluzionisti, con la loro tendenza a focalizzare l'attenzione esclusivamente su adattamenti immediati a condizioni locali, finiscono per ignorare i vincoli architettonici compiendo inversioni di spiegazione del tipo di quelle menzionate.

A titolo esemplificativo, riportiamo un caso recentemente dibattuto nella letteratura biologica sull'adattamento. L'antropologo Michael Harner (1977) ha formulato l'ipotesi che i sacrifici umani fra gli Aztechi potrebbero essere sorti come soluzione alla penuria cronica di carne (gli arti delle vittime venivano spesso mangiati, ma solo da persone di alto rango). E. O. Wilson (1978) ha poi inteso questa spiegazione come una fondamentale evidenza della predisposizione genetica adattiva dell'uomo ad essere carnivoro. Harner e Wilson ci chiedono insomma di considerare questo elaborato sistema sociale e il suo complesso insieme di giustificazioni, che includeva miti, simboli e tradizioni, come meri epifenomeni generati da una inconscia razionalizzazione della ragione “reale” di tutto ciò: il bisogno di proteine. Sahlins (1978), invece, ritiene che il sacrificio umano rappresentasse solo una parte del complicato meccanismo culturale degli Aztechi, che, nella sua interezza, non solo rappresentava l'espressione materiale della loro cosmologia, ma assolveva anche a certe funzioni pratiche, come la conservazione dei ranghi sociali e quella del sistema dei tributi fra le città.
Sospettiamo fortemente che il cannibalismo degli Aztechi fosse un “adattamento” tanto quanto lo sono gli evangelisti e i fiumi dei pennacchi, o i rosoni ornamentali delle volte a ventaglio: un epifenomeno secondario rappresenta un uso proficuo di parti disponibili, ma non la causa dell'intero sistema.
Diciamolo chiaramente: un sistema sviluppato per altre ragioni potrebbe aver generato una cospicua disponibilità di corpi freschi; così qualcuno poi potrebbe aver pensato di utilizzarli.
Perché invertire l'intero meccanismo e concepire paradossalmente tutta una cultura come epifenomeno di un'insolita maniera di procurarsi la carne?
I pennacchi non esistono per farvici dimorare gli evangelisti. (Inoltre, come lo stesso Sahlins afferma, non è affatto scontato che il sacrificio umano fosse un adattamento.) Le pratiche culturali umane possono essere ortogenetiche e condurre verso l'estinzione secondo modalità inaccessibili ai processi darwiniani, basati come sono sulla selezione genetica. Ad esempio, potrebbe essere che ciascun nuovo monarca volesse superare il suo predecessore in sacrifici sempre più elaborati e copiosi, cosicché la pratica finì per diventare di fatto un principio di saccheggiamento di risorse, fino al punto di rottura. Non sarebbe la prima volta che una cultura umana implode. Sono molti gli esperti che dubitano dell'assunto di partenza di Harner (Ortiz de Montellano, 1978).
Essi sostengono che fonti alternative di proteine non mancassero affatto; e una pratica che veicolava carne solo a persone privilegiate, le quali tra parentesi ne avevano già abbastanza, o che adoperava i corpi in modo così poco funzionale (si ricorda che solo gli arti erano consumati, e anche parzialmente), rappresenta di certo una strada poco o affatto lungimirante per aprire una macelleria.
Stiamo adoperando deliberatamente esempi non biologici, in una sequenza che va da casi alquanto remoti ad altri più familiari: dall'architettura all'antropologia. Facciamo questo perché l'importanza del vincolo strutturale o della natura epifenomenica dell'adattamento in essi non è offuscata dai nostri pregiudizi biologici. Si spera che il messaggio non venga sottovalutato dai biologi: se questi fossero stati degli esempi biologici, non avremmo forse considerato per forza d'abitudine l'adattamento epifenomenico come primario e cercato di imperniare un intero sistema strutturale su di esso?
Il bersaglio della nostra critica è un modo di pensare profondamente radicato tra gli studiosi dell'evoluzione. Lo denominiamo programma adattamentista o paradigma panglossiano. Esso è basato sull'idea (che non fu di Darwin, come vedremo), resa popolare da A. R. Wallace e A. Weismann verso la fine del XIX secolo, della quasi totale onnipotenza della selezione naturale nel forgiare le strutture organismiche e modellare il migliore dei mondi possibili.
Tale programma considera la selezione naturale tanto potente e non soggetta a limitazioni che la produzione diretta di adattamenti nel corso dell'intero suo processo diviene la causa primaria di pressoché tutte le forme, le funzioni e i comportamenti degli organismi. Certamente sono noti alcuni vincoli che si oppongono alla forza pervasiva della selezione naturale (tra questi soprattutto l'inerzia filetica, sebbene i vincoli strutturali immediati, come si dirà nell'ultimo paragrafo, raramente siano riconosciuti). Essi sono però di solito lasciati in disparte, perché considerati ininfluenti o per altri motivi; a volte, fatto ancor più triste, vengono semplicemente ammessi, evitando poi di trattarne seriamente o di chiamarli in causa.
Gli studi ispirati al programma adattamentista consistono in genere di due fasi.
(1) Innanzitutto si scinde un organismo in vari “tratti”, che vengono spiegati come strutture ottimamente modellate dalla selezione naturale per le loro funzioni. Per ragioni di brevità, non possiamo qui soffermarci sulla seguente questione fondamentale: cos'è un “tratto”? Alcuni evoluzionisti possono pensare che la cosa sia insignificante, o considerarla un problema puramente linguistico. Non è così. Gli organismi sono entità integrate, non collezioni di oggetti discreti. Come D'Arcy Thompson amava sottolineare (1942), gli evoluzionisti spesso sono stati portati fuori strada da riduzionismi inappropriati.
Il nostro esempio favorito concerne il mento umano (Gould 1977, pp. 381382; Lewontin 1978). Se consideriamo il mento come una “cosa”, piuttosto che come il prodotto dell'interazione fra due campi di crescita (alveolare e mandibolare), allora siamo portati a interpretare la sua origine in una maniera (ricapitolazione) esattamente opposta a quella che viene oggi generalmente accettata (neotenia).
(2) Se il tentativo di ottimizzazione parte dopo parte fallisce, scatta allora la regola dell'interazione tra di esse, attraverso il riconoscimento del principio che un organismo non può ottimizzare ciascuna parte senza imporre dei costi ad altre. Viene quindi introdotta la nozione di “compromesso”, e gli organismi sono visti come il miglior compromesso
possibile tra differenti domande selettive. In questo modo, l'interazione fra le parti viene completamente racchiusa all'interno del programma adattamentista. Qualunque imperfezione di una parte può essere spiegata nei termini del suo contributo alla migliore strutturazione possibile del tutto. Di solito non si pensa affatto che l'imperfezione sia il risultato di qualcosa di diverso rispetto all'azione immediata della selezione naturale.
Il Dr. Pangloss, spiegando a Candido perché egli soffrisse della malattia venerea, diceva: «È indispensabile nel migliore dei mondi, cioè proprio nel nostro. Poiché se Colombo, visitando le Indie dell'Ovest non avesse contratto il male, che avvelena la fonte della generazione, frequentemente la impedisce ed è con ciò chiaramente opposto al grande scopo della Natura, noi non avremmo il cioccolato né il rosso carminio».
Il programma adattamentista è veramente panglossiano. Il nostro mondo può non essere buono in tutto e per tutto, ma è il migliore che si possa avere. Ciascun tratto gioca la sua parte, e deve essere così com'è.
A questo punto alcuni evoluzionisti contesteranno il fatto che stiamo ridicolizzando la loro concezione dell'adattamento. Dopo tutto, essi non ammettono anche la deriva genetica, l'allometria e una varietà di altri casi di evoluzione non adattiva? Lo fanno, certo, ma qui si pone una questione differente. Nella storia naturale, tutte le cose possibili accadono di tanto in tanto; dunque, si dovrebbe evitare di avvalorare l'ipotesi preferita dichiarando l'impossibilità teorica di quelle rivali. Generalmente, si riconosce loro una qualche fondatezza, ma se ne delimita il campo d'azione così tanto da non interferire negli affari della natura. Poi ci si congratula con se stessi per essere ecumenici e non dogmatici.
Noi affermiamo che la maniera di argomentare appena illustrata abbia in genere relegato nell'ambito dell'irrilevante ogni alternativa alla logica della selezione del miglior modello possibile. Non abbiamo forse sentito tutti il catechismo sulla deriva genetica? Essa è importante solo in popolazioni così piccole da rischiare di estinguersi ancor prima di svolgere un qualche ruolo significativo nell'evoluzione (eppure vedi Lande 1976).
Dal riconoscimento di principio dell'esistenza di alternative non discende che esse vengano prese poi seriamente in considerazione nella pratica ordinaria. Sappiamo tutti che non ogni cosa è adattiva; tuttavia, di fronte ad un organismo, si tende a scomporlo in parti e a raccontare storie adattive come se i compromessi tra parti antagoniste, e ben disegnate, fossero la sola limitazione alla perfezione di ciascun tratto. È una vecchia abitudine.
Ecco come Romanes ebbe a dolersi di A. R. Wallace nel 1900: «Il signor Wallace non sostiene espressamente l'impossibilità teorica di leggi e cause altre rispetto a quelle dell'utilità e della selezione naturale. [...] Nondimeno, egli non riconosce in alcun luogo qualche altra legge o causa [...], egli praticamente conclude che, su basi empiriche o induttive, non c'è altra legge o causa da prendere in considerazione».
Il programma adattamentista può essere rintracciato diffusamente nella maniera di argomentare comune della biologia evoluzionistica. Qui di seguito ne diamo alcuni esempi, sperando che siano noti un po' a tutti.
(1) Se una giustificazione adattiva viene meno, cercane un'altra.
Le commessure a zigzag di bivalvi e brachiopodi, un tempo diffusamente considerate come un dispositivo per rinforzare la conchiglia, sono diventate nel corso del tempo dei setacci per filtrare particelle al di sopra di una data misura (Rudwick 1964). Una gran quantità di strutture esterne (aculei, corna, zanne), un tempo viste come armi contro i predatori, sono divenute elementi distintivi della competizione maschile intraspecifica (Davitashvili 1961). La faccia degli esquimesi, ritenuta un tempo “progettata per il freddo” (Coon et al. 1950), è divenuta un adattamento per generare e sostenere grandi forze masticatorie (Shea 1970). Non bisogna attaccare le interpretazioni più recenti; esse possono essere del tutto esatte. Ci meraviglieremmo, tuttavia, se il fallimento di una spiegazione adattiva debba sempre e comunque ispirare la ricerca di un'altra della medesima forma generale, piuttosto che considerare alternative all'idea che ciascuna parte serva per qualche scopo specifico.
(2) Se una giustificazione adattiva viene meno, ne esisterà un'altra.
(Versione più debole dell'argomento precedente.)
Costa e Bisol (1978), ad esempio, speravano di trovare una correlazione tra il polimorfismo genetico e la stabilità delle condizioni ambientali nel mare profondo, ma hanno fallito. La loro conclusione (1978; pp. 132133) è stata la seguente: «Il grado di polimorfismo genetico rilevato sembra non essere correlato a particolari fattori ambientali che caratterizzano l'area saggiata. I risultati suggeriscono che le strategie adattive di organismi appartenenti a phyla diversi sono differenti».
(3) In mancanza di una buona giustificazione adattiva, di primo acchito, attribuisci il fallimento all'imperfetta conoscenza dell'habitat e delle abitudini di vita dell'organismo.
Un altro vecchio argomento. Consideriamo ciò che pensava Wallace sul motivo per cui tutti i dettagli sul colore e sulla forma delle lumache di terra devono essere adattivi, anche se animali differenti abitano lo stesso ambiente (1899; p. 148): «L'esatta proporzione delle varie specie di piante, la quantità di ciascun gruppo di insetti o di uccelli, la peculiarità della maggiore o minore esposizione al sole o al vento in certi periodi critici e altre leggere differenze, per noi totalmente indifferenti o difficilmente riconoscibili, possono essere di grande rilevanza per queste umili creature e del tutto sufficienti per richiedere alcuni leggeri aggiustamenti nella dimensione, nella forma o nel colore a cui la selezione naturale porterà».
(4) Enfatizza l'utilità immediata ed escludi altri attributi di forma.
Verso la fine della didascalia che accompagna il Tyrannosaurus in vetroresina del Museo della Scienza di Boston troviamo scritto: «Il rompicapo delle zampe anteriori. In che modo il Tyrannosaurus adoperasse le sue minuscole zampe anteriori resta un mistero per la scienza. Esse sono troppo corte anche per arrivare alla bocca. Forse servivano per aiutare l'animale a rialzarsi da terra». (Abbiamo volutamente scelto un esempio basato sull'impatto pubblico della scienza, per mostrare come le abitudini del programma adattamentista si estendano ampiamente. Non vogliamo adoperare gli animali in vetroresina come uomini di paglia; il fatto è che argomenti simili, con un'enfasi espressa certo mediante parole differenti, compaiono regolarmente nella letteratura professionale.)
Non dubitiamo del fatto che il Tyrannosaurus adoperasse le sue minuscole zampe anteriori per fare qualcosa; e se fossero sorte ex novo, nel corso dell'evoluzione, avremmo magari anche incoraggiato la ricerca di una qualche ragione adattiva. Il punto è che esse sembrano però essere semplicemente il prodotto di una riduzione degli arti omologhi dei loro antenati, ritenuti generalmente funzionali (per esempio, i lunghi arti dell'allosauro). Così stando le cose, forse non c'è alcun bisogno di una spiegazione adattiva diretta della loro riduzione. È probabile che fosse uno sviluppo correlato di campi allometrici per incremento relativo nelle dimensioni del capo e delle zampe posteriori. Questa ipotesi non adattiva può essere testata mediante metodi allometrici convenzionali (Gould 1974, in generale; Lande 1978, sulla riduzione degli arti), e ci pare più interessante e feconda di speculazioni non testabili basate su un qualche tipo di utilizzazione secondaria nel migliore dei mondi possibili.
Non bisogna confondere il fatto che una struttura venga in qualche maniera e misura utilizzata (si pensi ai pennacchi, agli spazi nelle volte a ventaglio o ai corpi degli Aztechi) con la ragione evolutiva primaria della sua esistenza e conformazione.
«Tutto questo è una manifestazione della giustezza delle cose, perché se ci fosse un vulcano a Lisbona, non potrebbe essere da alcuna altra parte. Perciò è impossibile per le cose non essere dove sono, perché ogni cosa è per il meglio». (Commento del Dr. Pangloss al grande terremoto di Lisbona del 1755, in cui persero la vita circa 50.000 persone.)
Non ci opporremmo così strenuamente al programma adattamentista se la sua invocazione, in ciascun caso specifico, non conducesse in linea di principio al rifiuto dell'evidenza. Potremmo anche considerarlo semplicemente limitato e criticare soltanto la sua condizione di argomento di prima scelta. L'importante è che ogni qualvolta esso dovesse venir meno, per il fallimento di qualche verifica esplicita, allora le alternative ad esso possibili dovrebbero avere una loro chance.
Sfortunatamente, però, procedure e modi di fare comuni tra gli evoluzionisti non consentono affatto una simile rinuncia di principio, e questo per almeno due ragioni. In primo luogo, la rinuncia ad una storia adattiva di solito conduce alla sua sostituzione con un'altra, senza neanche farsi sfiorare dall'idea che possa essere più opportuno pensare a un differente tipo di spiegazione. E siccome la quantità di storie adattive possibili è grande tanto quanto le nostre menti sono fertili, è sempre possibile costruire nuove storie. Se una storia non è disponibile nell'immediato, ci si può sempre appellare a una temporanea ignoranza e aver fiducia che prima o poi qualcosa arriverà (proprio come accadeva nell'esempio di Costa e Bisol, prima menzionato).
In secondo luogo, i criteri per l'accettazione di una storia adattiva sono così vaghi e nebulosi che molte ipotesi passano senza conferma alcuna. Spesso gli evoluzionisti adoperano la coerenza rispetto al meccanismo della selezione naturale come criterio unico di discernimento, considerando il loro lavoro compiuto quando abbiano escogitato una storia plausibile, e questo è praticamente sempre possibile. Per avviare una ricerca storica, in fondo, basta definire opportuni criteri di scelta di spiegazioni proprie all'interno della copiosa serie di percorsi di sviluppo possibili di ogni odierno risultato.
Uno di noi (Gould 1978), per esempio, ha criticato il lavoro di Barash sull'aggressione degli uccelli azzurri di montagna proprio per questo motivo. Barash ha montato un maschio impagliato vicino ai nidi di due coppie di uccelli azzurri mentre il maschio era fuori in cerca di cibo. Ha ripetuto l'esperienza con gli stessi nidi per tre volte, a intervalli di dieci giorni: una volta prima della deposizione delle uova, le altre due dopo. Sono stati quindi contati gli attacchi fatti dal maschio, sia verso lo zimbello che verso le femmine. La prima volta ci sono stati attacchi violenti in entrambi i nidi, rivolti più verso lo zimbello che verso le femmine. L'aggressività verso lo zimbello è poi diminuita fortemente nella seconda e terza volta, e quella verso le femmine è scesa pressoché a zero. Barash ha concluso che tutto ciò ha un significato evoluzionistico, perché i maschi si mostrano più sensibili alla presenza di intrusi prima che le uova siano deposte che dopo la deposizione (quando essi possono avere ormai una certa sicurezza che i loro geni siano già dentro). Avendo escogitato questa storia plausibile, egli ha considerato compiuto il suo lavoro (1976, pp. 1099-1100).
I risultati concordano con le aspettative della teoria evoluzionistica. È chiaro che l'aggressione verso il maschio intruso (lo zimbello) dovrebbe essere particolarmente vantaggiosa prima della stagione riproduttiva, quando in genere si lotta per i territori o i nidi, e si difendono. [...] Anche la risposta aggressiva iniziale nei confronti della compagna è adattiva, poiché data la situazione di un possibile tradimento (i.e., la presenza dello zimbello maschio vicino alla femmina), pur assumendo che femmine fertili sostitutive siano comunque disponibili, la conquista di una nuova compagna da parte dell'altro maschio aumenterebbe appunto la fitness dell'altro a discapito della propria. [...] La diminuzione dell'aggressività maschiofemmina durante l'incubazione e dopo la schiusa delle uova, a sua volta, potrebbe invece attribuirsi al fatto di non poter essere più traditi. [...] Come si vede, i risultati concordano con una spiegazione di tipo evoluzionistico.
Tutto questo ragionamento potrebbe essere giusto, ma cosa dire di una spiegazione alternativa alquanto banale che Barash liquida senza alcun test? Ecco: quando il maschio ritorna la seconda e la terza volta incontra lo stesso zimbello, lo saggia un po' e lo riconosce come lo stesso falso che aveva visto in precedenza, che non infastidisce più di tanto le sue femmine. Barash non ha eseguito alcuna verifica di questa spiegazione alternativa alla sua storia adattiva convenzionale. Ad esempio, poteva mostrare a un maschio lo zimbello per la prima volta dopo che le uova erano state deposte.
Poiché il lavoro di Barash è stato criticato, Morton et al. (1978) hanno ripetuto l'esperienza, con una piccola variazione (cioè l'intromissione anche di zimbelli femmina), adoperando l'uccelloazzurro orientale Sialia sialis, strettamente imparentato al precedente. «Speravamo di confermare – essi hanno poi scritto – che le conclusioni di Barash rappresentassero un fatto concreto su base eminentemente evoluzionistica, almeno all'interno del genere Sialia. Purtroppo, non possiamo farlo». Non sono stati affatto riscontrati comportamenti “antitradimento”: i maschi non si rivolgevano alle loro femmine aggressivamente, dopo aver testato lo zimbello, in nessuno stadio del nido. Al contrario, le femmine spesso cercavano di approcciare lo zimbello maschio e, d'altronde, attaccavano comunque lo zimbello femmina più di quanto i maschi attaccassero i loro. «Questa risposta violenta risultava pressoché prossima alla distruzione dello zimbello femmina, nel corso dei primi incontri, arrivando fino alla sua completa distruzione nei successivi, con una delle femmine che volava via con la testa dell'uccello impagliato, staccata abbastanza presto nel corso dell'esperimento e lasciata poi cadere giù nel sottobosco» (1978, p. 969).
Non potendo ricorrere alla spiegazione su base selettiva di Barash, però, a questo punto, essi ne hanno escogitato una tutta propria, che renda conto di entrambi i risultati alla maniera adattamentista. Forse, hanno pensato, le femmine sostitutive sono scarse nella specie in questione e abbondanti nella specie con cui ha lavorato Barash, e ciò influisce sul comportamento aggressivo dei maschi verso le femmine. Poiché i maschi di Barash possono facilmente sostituire una femmina “infedele” con un'altra, essi si permettono di essere con loro schizzinosi e possessivi. I maschi dell'uccelloazzurro orientale, invece, hanno compagne poco numerose e probabilmente è meglio rispettarle. Essi quindi concludono: «Pur non appoggiando l'idea di Barash che i maschi degli uccelliazzurri abbiano adattamenti antitradimento, noi suggeriamo che entrambi gli studi conducano a “risultati [che] concordano con le aspettative della teoria evoluzionistica” (Barash 1976, p. 1099), come avviene nel caso di ciascuno studio che possa dirsi accurato».
Come deve sembrare buona una teoria da cui non possono provenire studi che non siano accurati (con l'espressione “teoria evoluzionistica”, infatti, essi intendono chiaramente l'azione della selezione naturale applicata a casi particolari, piuttosto che il fenomeno del cambiamento in sé).
Poiché Darwin ha ottenuto il saio del santo (se non la divinità) tra gli evoluzionisti, e poiché da tutte le parti si invoca la fedeltà a Dio, egli è stato spesso dipinto come un selezionista radicale, in cuor suo, che faceva ricorso ad altri meccanismi solo per ripiego o se costretto dalla compianta ignoranza della sua epoca in materia di ereditarietà. Questo punto di vista è falso. Sebbene Darwin considerasse la selezione come il più importante dei meccanismi evolutivi (come facciamo del resto anche noi), nessun argomento dei suoi oppositori lo faceva più irritare del comune tentativo di caricaturizzare o banalizzare la sua teoria affermando che essa fosse basata esclusivamente sulla selezione naturale. Nell'ultima edizione de L'origine delle specie, egli scriveva (1872, p. 395):
Poiché le mie conclusioni sono state recentemente travisate, e si è detto che io attribuisco la modificazione delle specie esclusivamente alla selezione naturale, mi permetto di sottolineare che nella prima edizione di questo lavoro, e anche successivamente, ho collocato in una posizione preminente – cioè alla fine dell'Introduzione – le seguenti parole: «Sono convinto che la selezione naturale sia stato il principale, ma non esclusivo, mezzo di modificazione». Ciò non è stato di alcuna utilità. La portata del costante fraintendimento è ancora grande.
Romanes, il cui primo famoso saggio sul pluralismo di Darwin (1900) contro il panselezionismo di Wallace e Weismann meriterebbe una diversa considerazione, commentava le osservazioni di Darwin appena menzionate come segue: «Nell'intera serie degli scritti di Darwin non può trovarsi una sola affermazione così energicamente formulata; essa rappresenta la sola nota di amarezza fra tutte le migliaia di pagine che egli ha pubblicato». E a quanto sembra, Romanes non conosceva la lettera che Darwin scrisse a Nature nel 1880, in cui rimproverava a Sir Wyville Thomson di aver caricaturizzato la sua teoria come panselezionista:
Sono dispiaciuto di scoprire che Sir Wyville Thomson non ha compreso il principio della selezione naturale. [...] Se così non fosse, egli non avrebbe certo scritto la seguente affermazione nell'Introduzione del suo libro intitolato Il viaggio del Challenger: «Il carattere della fauna abissale non accorda alcun sostegno alla teoria che attribuisce l'evoluzione delle specie all'esistenza di una grande variabilità guidata dalla sola selezione naturale». Questo è un genere di polemica non insolito per teologi e metafisici quando scrivono di argomenti scientifici, ma è di certo una novità che provenga da un naturalista. [...] Può Sir Wyville Thomson citare qualcuno che abbia mai affermato che l'evoluzione delle specie dipende dalla sola selezione naturale? Per quanto mi riguarda, credo che nessuno abbia fornito così tante osservazioni sugli effetti dell'uso e del non uso delle parti come ho fatto io nel lavoro intitolato Variazione degli animali e delle piante allo stato domestico; e tali osservazioni furono svolte proprio su questo speciale argomento. Ho addotto in proposito anche un considerevole corpo di fatti che mostra l'azione diretta delle condizioni esterne sugli organismi.
Ovviamente, oggi non è possibile ritenere che tutti i meccanismi sussidiari di Darwin debbano essere considerati sensati e validi, sebbene molti di essi, includendo la modificazione diretta e le correlazioni di crescita, siano in realtà molto importanti. Bisogna tuttavia apprezzare la sua costante attitudine al pluralismo nel tentare di spiegare la complessità della Natura.
Nello spirito pluralistico proprio di Darwin, presentiamo qui di seguito una serie incompleta di possibili alternative all'adattamento immediato, per la spiegazione di forme, funzioni e comportamenti.
(1) Nessun adattamento e nessuna selezione affatto.
Attualmente i genetisti di popolazione sono in netto disaccordo sulle modalità con cui la maggior parte del polimorfismo genetico all'interno delle popolazioni e altre differenze genetiche tra le specie possano essere di fatto il risultato di una selezione naturale, in opposizione a fattori puramente casuali. Le popolazioni sono in genere limitate nelle dimensioni e quelle isolate, che costituiscono il primo passo nel processo di speciazione, sono spesso costituite da un numero molto piccolo di individui. Come risultato di questa restrizione nelle dimensioni della popolazione, le frequenze degli alleli cambiano mediante deriva genetica, un tipo di errore genetico casuale saggiato. Il processo stocastico di cambiamento nella frequenza genica mediante deriva genetica casuale, che include il forte processo di campionamento che avanza quando una nuova popolazione isolata si forma da pochi individui emigrati, ha svariate importanti conseguenze.
In primo luogo, le popolazioni e le specie possono divenire geneticamente differenziate, magari anche con la fissazione di alleli differenti in uno stesso locus, nella completa assenza di qualsivoglia forza selettiva.
In secondo luogo, gli alleli possono fissarsi in una popolazione anche a dispetto della selezione naturale. Sebbene un gene sia favorito dalla selezione naturale, infatti, qualche porzione della popolazione, che dipende dal prodotto della grandezza della popolazione N per l'intensità della selezione s, potrà divenire omozigote per un allele meno vantaggioso a causa della deriva genetica. Se Ns è ampio, questa fissazione casuale di alleli non favorevoli è un fenomeno raro, ma se i coefficienti di selezione sono dell'ordine del reciproco della grandezza della popolazione (Ns = 1) o più piccoli, la fissazione di alleli svantaggiosi è alquanto comune. Se sono molti i geni coinvolti nella determinazione di un carattere metrico come la sagoma, il metabolismo o il comportamento, allora l'intensità della selezione in ciascun locus sarà piccolo e anche il prodotto Ns per locus lo sarà di conseguenza. Come risultato, in molti loci andranno a fissarsi alleli non ottimali.
In terzo luogo, le nuove mutazioni hanno una piccola chance di essere incorporate in una popolazione, anche quando selettivamente favorevoli. La deriva genetica causa l'immediata perdita di molte nuove mutazioni subito dopo la loro introduzione. Con un'intensità di selezione s, una nuova mutazione favorevole ha una probabilità di appena 2s di essere incorporata. Così stando le cose, è praticamente impossibile affermare che una nuova mutazione che torni utile per qualche ragione adattiva possa eventualmente diffondersi. Si tratta di un tempo estremamente lungo se si pensa che una sola mutazione su 1000 o una su 10000 può rivelarsi “opportuna” ed essere magari poi anche incorporata in una popolazione.
(2) Nessun adattamento e nessuna selezione sulla parte in questione: la forma della parte è una conseguenza correlata della selezione diretta altrove.
Sotto questa importante categoria, Darwin classificava le sue “misteriose” leggi della “correlazione di crescita”. Oggi parliamo di pleiotropia, allometria, “compensazione materiale” (Rensch 1959, p. 1791-87) e correlazioni meccaniche forzate, nel senso di D'Arcy Thompson (1942; Gould 1971). Qui tocchiamo con mano l'idea di organismo come un tutt'uno integrato, essenzialmente non scomponibile in parti indipendenti e ottimizzabili separatamente.
Sebbene i modelli allometrici di sviluppo siano soggetti a selezione come lo è la morfologia statica in sé (Gould 1966), alcune regolarità nella crescita relativa probabilmente non sono sotto un diretto controllo adattivo. Per esempio, non dubitiamo del fatto che la ben nota allometria interspecifica di 0.66 nelle dimensioni del cervello in tutti i maggiori gruppi di vertebrati rappresenti un “criterio di organizzazione” selezionato, tuttavia la sua importanza rimane incerta (Jerison 1973). Il dato può essere ottenuto in un intervallo tassonomico troppo ampio e dice semplicemente che tutte le creature considerate sono in genere ben proporzionate nelle dimensioni relative del loro corpo. Tuttavia un'altra comune allometria, la scala intraspecifica 0 concernente la taglia corporea di adulti omeotermi che differiscono tra loro per dimensioni fisiche, o appartenenti a razze diverse all'interno di una stessa specie, non richiede probabilmente una storia selettiva, anche se molti, incluso uno di noi (Gould 1974), hanno cercato di fornirne una. R. Lande (in una comunicazione personale) ha adoperato l'esperienza di Falconer (1973) per mostrare che la selezione sulle sole dimensioni del corpo porta nel topo a un rapporto cervello/corpo di 0.35 attraverso poche generazioni.
Altri esempi, numerosi e forse anche maggiormente significativi, si trovano nella letteratura concernente le alterazioni dell'intervallo di tempo della maturazione sessuale (Gould 1977). Almeno tre volte nell'evoluzione degli artropodi (acari, mosche e scarafaggi), uno stesso complesso adattamento pare si sia evoluto per un rapida alternanza generazionale in animali fortemente selezionati mangiatori di funghi abbondanti ma effimeri: le femmine si riproducono quando sono ancora larve e crescono portando la successiva generazione all'interno dei loro stessi corpi. La prole divora le madri dall'interno ed emerge poi dall'involucro ormai cavo per essere divorata pochi giorni dopo dalla successiva progenie. In questo caso, sarebbe assurdo cercare un significato adattivo nella morfologia pedomorfica in sé, perché si tratta invece di un sottoprodotto della selezione di cicli più rapidi nel corso delle generazioni. In alcuni casi ancor più interessanti, una selezione a favore di piccole dimensioni (come tra gli animali della fauna interstiziale) o una rapida maturazione sessuale (come nei maschi nani di molti crostacei) è avvenuta mediante progenesi (Gould 1977, pp. 324336), e gli adulti discendenti possiedono un misto di caratteristiche ancestrali adulte e giovanili. Molti biologi sono stati tentati di cercare un significato adattivo primario per questa strana commistione, tuttavia essa probabilmente origina come sottoprodotto dell'arresto della maturazione, che lascia “ferme” alcune caratteristiche allo stadio larvale, mentre permette che altre, più fortemente correlate alla maturazione sessuale, raggiungano la configurazione adulta dei loro antenati.
(3) Riaccoppiamento di selezione e adattamento.
(3a) Selezione senza adattamento.
Lewontin (1979) ha presentato il seguente esempio ipotetico.
Una mutazione che raddoppia la fecondità individuale può far aumentare rapidamente le dimensioni di una popolazione. Se non avviene anche un cambiamento di efficienza nell'utilizzazione delle risorse, tuttavia, gli individui non lasceranno più prole di prima: essi semplicemente deporranno un numero maggiore di uova, ma la prole in eccesso morirà per la limitatezza delle risorse. In che senso si può dire che gli individui o la popolazione sono meglio adattati che in precedenza? Se un predatore di uova o di stadi immaturi si interessasse alla specie in questione proprio perché sia le une che gli altri sono ora più abbondanti, le dimensioni della popolazione potrebbero in realtà addirittura diminuire. Ciò nonostante, la selezione naturale favorirà tutte le volte gli individui con maggiore fecondità.
(3b) Adattamento senza selezione.
Molti organismi marini sessili, tra cui soprattutto spugne e coralli, sono molto ben adattati al regime di correnti d'acqua in cui essi vivono. Ma numerosi “buoni piani di organizzazione” possono avere un'origine meramente fenotipica, indotta per lo più dalla corrente stessa. (Possiamo essere sicuri di ciò in numerosi casi; ad esempio, quando individui geneticamente identici di una colonia assumono forme diverse in microhabitat differenti.) Modelli più estesi di variazione geografica sono spesso “adattivi”, ma su base meramente fenotipica. Sweeney e Vannote (1978), per esempio, hanno mostrato che molti insetti acquatici emimetaboli da adulti presentano dimensioni piccolissime e una ridotta fecondità quando crescono a temperature al di sotto o al di sopra di quella per loro ottimale. Coerentemente, i modelli di correlazione climatica nella distribuzione geografica di questi insetti – così spesso assunti a priori come chiari segni di adattamento genetico – potrebbero semplicemente riflettere questa plasticità fenotipica.
Ciò che è detto “adattamento” – cioè l'opportuno adeguamento degli organismi al proprio ambiente – può avvenire su tre livelli gerarchici e con cause differenti. È una sfortuna che il nostro linguaggio abbia fatto convergere tutto verso un unico risultato, chiamando tutti e tre i fenomeni con il nome di “adattamento”: in tal modo, le differenze nei processi sono state oscurate e gli evoluzionisti sono stati spesso fuorviati, estendendo la modalità darwiniana agli altri due livelli. In primo luogo, abbiamo ciò che i fisiologi dicono “adattamento”: la plasticità fenotipica che permette agli organismi di forgiare la loro forma secondo le circostanze prevalenti nel corso della loro ontogenesi. Gli “adattamenti” umani alle grandi altitudini rientrano in questa categoria (mentre altri, come la resistenza alla malaria degli anemici falciformi in eterozigosi, sono genetici o darwiniani).
Gli adattamenti fisiologici non sono ereditabili, sebbene lo sia presumibilmente la capacità di svilupparli. In secondo luogo, abbiamo una forma “ereditabile” di adattamento nondarwiniano negli uomini (e, in maniera rudimentale, in poche altre specie sociali avanzate): l'adattamento culturale (trasmesso mediante l'insegnamento). Molti confusi pensieri della sociobiologia umana sorgono da una mancata distinzione tra questa modalità di adattamento e quella di tipo darwiniano basato sulla variazione genetica.
In terzo luogo, abbiamo l'adattamento generato mediante il meccanismo convenzionale darwiniano della selezione esercitata sulla variazione genetica. La sola osservazione di una opportuna adeguatezza di un organismo al suo ambiente non è affatto sufficiente per inferire l'azione della selezione naturale.
(4) Adattamento e selezione, ma nessuna base selettiva per le differenze fra gli adattamenti.
Specie di organismi tra loro collegati, o differenti popolazioni all'interno di una specie, sviluppano spesso adattamenti diversi come soluzione di un medesimo problema. Quando i “picchi adattivi multipli” sono tutti occupati, non abbiamo alcuna base per asserire che una soluzione sia migliore di un'altra. La soluzione seguita in ogni passaggio filogenetico è un risultato di natura storica: i primi passi procedono in una certa direzione, vantaggiosa, ma altre linee di sviluppo potrebbero condurre verso una prosperità altrettanto adeguata. Ogni naturalista possiede la sua esemplificazione favorita sull'argomento.
Nell'India occidentale, ad esempio, le popolazioni della chiocciola Cerion vivono su coste rocciose e ventilate, sviluppando quasi sempre gusci bianchi, spessi e relativamente tozzi, per note ragioni adattive. Possiamo identificare almeno due diversi percorsi di formazione del colore bianco a partire dalla chiazza che si forma nei primi giri di tutte le Cerion, due differenti modalità di ispessimento del guscio calcareo e tre stili allometrici di sviluppo delle conchiglie tozze. Queste dodici possibili combinazioni possono essere tutte individuate e riconosciute nelle popolazioni bahamiane della specie, ma, ragionando esclusivamente in termini di piani ottimali di organizzazione piuttosto che di contingenza storica, sarebbe poi così sensato chiedersi perché le Cerion orientali di Long Island hanno evoluto una soluzione e quelle di Acklins Island un'altra?
(5) Adattamento e selezione, ma l'adattamento è una utilizzazione secondaria di parti presenti per ragioni strutturali, di sviluppo o storiche.
Abbiamo già discusso di questo argomento nella prima sezione del presente lavoro, trattando dei pennacchi, degli spazi mediani nelle volte a ventaglio e del cannibalismo. Se tra gli uomini la capacità di arrossire può essere considerata come un adattamento in fatto di selezione sessuale, questo non ci aiuta a comprendere perché il sangue è rosso. L'utilità immediata di una struttura organismica spesso non dice niente sulla ragione del suo venire in essere.
Nell'Europa continentale, gli evoluzionisti non sono mai stati molto attratti dal penchant angloamericano di scomporre gli organismi in parti distinte per poi cercare di spiegare ciascuna di esse come un adattamento diretto. Un'alternativa generale esiste, sia in una forma forte che debole. Nella forma forte, che è possibile riscontrare anche in teorici di grande importanza come Schindewolf (1950), Remane (1971) e Grassé (1977), la selezione naturale, del tipo di quella adoperata nel programma adattamentista, può spiegare le sole modificazioni superficiali del Bauplan, che adeguano la struttura all'ambiente: per esempio, le talpe sono cieche, le giraffe hanno il collo lungo e le anatre i piedi palmati. Ma i più importanti passi dell'evoluzione, la costruzione stessa del Bauplan e la transizione tra differenti Baupläne devono coinvolgere un qualche altro meccanismo sconosciuto, forse “interno”. Crediamo che i biologi inglesi abbiano fatto bene a rigettare questa forma forte, poiché conduce a una sorta di misticismo.
L'argomento ha comunque una forma debole – e paradossalmente potente – che non è stata apprezzata, ma merita di esserlo. Essa, come nel caso precedente, ammette la selezione di modificazioni superficiali del Bauplan e nega che il programma adattamentista (atomizzazione e selezione ottimizzante delle parti) possa riuscire a giustificare i Baupläne e la transizione tra loro. A questo punto, tuttavia, non c'è alcun ricorso a meccanismi sconosciuti. L'idea è che i piani di base del corpo degli organismi siano così integrati e pieni di vincoli adattivi (categorie 2 e 5 della nostra tipologia) che gli argomenti di tipo tradizionale a favore della selezione riescono a spiegare poco o niente intorno ad essi. Non si nega che il cambiamento, quando esso avvenga, possa essere mediato dalla selezione naturale, ma si ritiene che numerose costrizioni riducano il numero dei possibili percorsi e modi di cambiamento tanto fortemente che i vincoli stessi divengono di gran lunga l'aspetto più interessante dell'evoluzione.
Rupert Riedl (1975), lo zoologo austriaco che ha compendiato questa tesi per il pubblico inglese (1977), scrive:
Il mondo dei viventi sembra essere affollato di modelli universali di organizzazione che molto probabilmente non trovano una diretta spiegazione nelle condizioni ambientali o nelle radiazioni adattive, ma esistono primariamente per esigenze universali che possono essere soddisfatte solo sotto le condizioni sistemiche dell'organizzazione complessa in sé. [...] A causa dell'enorme e profondo pensiero collezionato nel campo della morfologia da Goethe a Remane, però, tutto ciò non risulta autoevidente, anzi è stato virtualmente tagliato fuori dalla biologia moderna. In molte università americane non si insegna affatto. E anche gli insegnanti che potrebbero insegnarlo sono scomparsi.(Rupert Riedl, Order in Living Systems. A Systems Analysis of Evolution, John Wiley and Sons, New York 1978; traduzione inglese a cura di R.P.S. Jefferies di Die Ordnung des Lebendigen, 1975).
I vincoli concernenti il cambiamento evolutivo possono essere suddivisi in almeno due categorie. A tutti gli evoluzionisti sono familiari i vincoli legati alla filogenesi, incorporati nella classica distinzione di Gregory (1936) tra abitudine ed eredità. Ammettiamo un tipo di inerzia filetica quando diciamo, ad esempio, che gli uomini non sono progettati ottimamente per la posizione eretta, perché gran parte del nostro Bauplan fu sviluppato per la vita da quadrupedi. Oppure invochiamo il vincolo filetico per spiegare perché nessun mollusco riesca a volare o nessun insetto sia grande quanto un elefante.
Abbiamo poi i vincoli legati allo sviluppo, che, pur essendo in realtà una sottocategoria dei vincoli filetici, possono render conto, almeno in teoria, di pressoché tutti i possibili percorsi evolutivi. Negli organismi complessi, i primi stadi dello sviluppo ontogenetico sono decisamente refrattari al cambiamento evolutivo, probabilmente perché la differenziazione di organi e sistemi, oppure la loro integrazione in un corpo funzionante sono processi molto delicati, facilmente degenerabili attraverso errori iniziali con effetto cumulativo. Le leggi fondamentali dell'embriologia di Von Baer (1828) rappresentano poco più che un semplice riconoscimento che i primi stadi dello sviluppo sono altamente conservativi, nonché fortemente refrattari a cambiamenti negli stadi successivi. E la legge biogenetica di Haeckel, cioè l'argomento principale dell'ultima biologia evoluzionistica del XIX secolo, giaceva su un fondamentale fraintendimento degli stessi dati (Gould 1977).
Se lo sviluppo ha a che fare con pacchetti integrati e non con pezzi sconnessi la cui evoluzione può essere seguita singolarmente, allora il programma adattamentista non può spiegare l'alterazione dei programmi di sviluppo sottostanti a quasi tutti i cambiamenti del Bauplan. Il paleontologo tedesco A. Seilacher, il cui lavoro meriterebbe molta più attenzione di quanta ne ha ricevuta, ha messo in evidenza i vincoli che egli chiama “bautechnischer” o architettonici (Seilacher 1970). Essi non derivano da precedenti adattamenti che vengono mantenuti in nuove collocazioni ecologiche (cioè i vincoli filetici generalmente riconosciuti), ma rappresentano dei limiti strutturali che non sono mai stati degli adattamenti quanto piuttosto la conseguenza necessaria di forme e materiali selezionati per costruire i Baupläne di base. Abbiamo dedicato la prima sezione di questo lavoro agli esempi non biologici di questa categoria. Qualora si decida di poggiare una cupola su archi a tutto sesto, i pennacchi esisteranno sin dal progetto. Qualcosa di analogo vale nel caso degli organismi. Su di essi i vincoli architettonici esercitano una vasta influenza. E l'argomento è pieno di intuizioni potenziali perché raramente è stato riconosciuto e trattato.
In un esempio affascinante, Seilacher (1972) ha mostrato che la forma divaricata nell'architettura dei gusci dei bivalvi (Figura 3) ricorre diffusamente in tutti i gruppi di molluschi e altrettanto nei brachiopodi. Questa forma di base si esprime in un'ampia varietà di strutture: linee ornamentali rialzate (che non vanno confuse con le linee di crescita, perché non si conformano al [p.596] margine del mantello neppure una volta), modelli di colorazione, strutture interne di mineralizzazione della calcite e solchi incisi. Seilacher non conosce ciò che genera questi modelli, ma sente che la tradizionale ed esclusiva insistenza sul valore adattivo di ciascuna determinazione possibile ha portato in secondo piano la questione della sua origine nel corso della crescita e ha impedito che il fenomeno venisse riconosciuto come un fatto generale. Esso deriva da qualche caratteristica disomogeneità nella crescita del mantello, che probabilmente produce modelli d'interferenza a partire da centri collocati in maniera regolare. Basta una semplice simulazione al computer per generare la forma in questo modo (Waddington e Cowe, 1969). Ed essendo il modello del tutto generale, non può essere affatto un adattamento diretto.
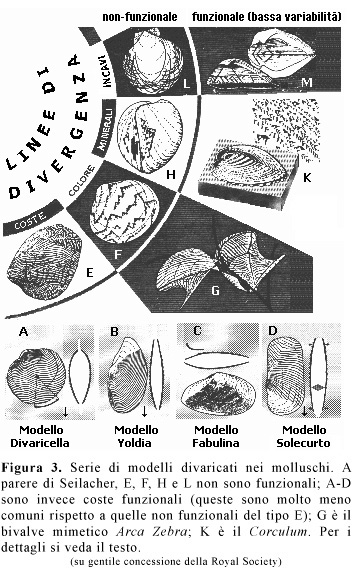
Seilacher sostiene che la maggior parte delle determinazioni del modello probabilmente non sono affatto adattive. Le ragioni di ciò sono varie, ma nel loro insieme assomigliano alle nostre. Alcune sono basate su osservazioni sul campo: alcuni modelli colorati, ad esempio, non servono a niente perché i bivalvi che li possiedono vivono generalmente infossati nei sedimenti marini, oppure sono ricoperti da un periostraco così spesso che i colori non si vedono affatto. Altre ragioni riguardano invece principi più generali, come ad esempio la presenza di certe caratteristiche solo in individui strani o patologici, la rarità come anomalia nello sviluppo, oppure la grande variabilità riscontrata nel caso di strutture in cui, essendo considerate funzionali su base ingegneristica, non dovrebbe esserci.
Solo in una ristretta minoranza di casi il modello divaricato risulta funzionale, come viene mostrato nelle quattro categorie della Figura 3. Le coste divaricate possono agire come palette e punti di ancoraggio per scavare (Stanley, 1970), sebbene sia chiaro che non sono sorte per una tale funzione. Quelle colorate sono mimetiche in una specie (Pteria zebra) che vive sulle ramificazioni dell'esoscheletro degli idrozoi; in questo caso infatti la variabilità è fortemente ridotta. La mineralizzazione delle coste ha natura sicuramente adattiva in una sola creatura, il caratteristico bivalve Corculum cardissa (in altre specie il fenomeno fa la sua comparsa in esemplari anomali, o risulta essere un prodotto postmortem dell'erosione della conchiglia). Questo bivalve è compresso unicamente in senso anteroposteriore e giace sul substrato con la parte posteriore in alto. Distribuiti sulla porzione di superficie esposta si trovano triangoli di mineralizzazione. Essi sono translucidi, mentre il resto della conchiglia è opaco. Sotto queste finestre dimorano alghe endosimbionti!
La letteratura tradizionale sulla struttura divaricata nei bivalvi ha focalizzato l'attenzione sul suo possibile significato adattivo (e ha spesso fallito). Ma Seilacher ha probabilmente ragione nel presentare questo caso in maniera analoga a quella in cui noi qui abbiamo trattato di pennacchi, di rosoni ornamentali e di corpi sacrificali, nel primo paragrafo. Il modello divaricato è fondamentalmente un vincolo architettonico. Solo occasionalmente, data la sua esistenza, è stato adoperato in vista di un qualche risultato vantaggioso. Di certo non è possibile comprenderne l'origine né il significato evolutivo considerando adattamenti infrequenti e secondari come il motivo principale dell'esistenza stessa del modello.
In un aneddoto sulle impronte digitali che ha come protagonista Herbert Spencer, Galton (1909, p. 257) criticava il programma adattamentista focalizzando l'attenzione sui vincoli e sui modi dello sviluppo:
Sulle creste papillari dei polpastrelli umani si è scritto tantissimo, ma l'ultima parola non è ancora stata pronunciata. In un uomo, in un dito esse formano spirali, mentre in altri nodi. Citerò in proposito un singolare aneddoto di Herbert Spencer. Egli mi chiese di mostrargli il mio laboratorio e di prendere le sue impronte, come poi feci. Gli parlai del fallimento dei tentativi di scoprire l'origine di queste strutture e di come le dita di bambini non ancora nati fossero state sezionate per conoscere i loro primi stadi di sviluppo, e così via. Spencer osservò che forse avevo cominciato nella maniera sbagliata, poiché dovevo dapprima considerare la funzione a cui le creste adempiono e poi lavorare a ritroso. Quanto allo scopo, egli disse, era ovvio che le delicate labbra delle ghiandole sudoripare richiedevano la protezione delle creste su ciascuna parte di esse, e a partire da ciò egli venne elaborando una consistente ipotesi di grande ingegno. Io risposi che le sue argomentazioni erano belle e meritavano di essere vere, ma in effetti le labbra dei dotti non corrono nelle valli tra le creste, bensì lungo le creste delle papille stesse.
Riteniamo che i vantaggi derivanti dall'abbandono del programma adattamentista siano davvero molto grandi. Non offriamo un consiglio disperato, come gli adattamentisti ci accusano di fare, perché non adattivo non significa non intelligibile. Diamo invece il benvenuto a tutta quanta la ricchezza che un approccio pluralistico può offrire, sentendoci con ciò vicini allo spirito di Darwin. Sotto il programma adattamentista, i grandi temi di natura storica legati alla morfologia dello sviluppo e al Bauplan di base sono stati largamente abbandonati; in effetti, se la selezione può rompere ogni correlazione tra le parti di un organismo e ottimizzare ciascuna di esse separatamente, la loro integrazione conta poco o niente.
Troppo spesso il programma adattamentista ci ha fornito una biologia evoluzionistica fatta di parti di organismi e di geni, ma non di organismi interi. Assumendo che ogni possibile transizione possa avvenire autonomamente, si sottovaluta l'importanza dei blocchi derivanti dallo sviluppo integrato e quella di altri vincoli di natura storica o strutturale. Una prospettiva pluralistica può invece far rientrare nuovamente gli organismi, con tutta la loro recalcitrante e tuttavia intelligibile complessità, nella teoria evoluzionistica.
Baer, K. E. von 1828 Entwicklungsgeschichte der Tiere. Königsberg: Bornträger.
Barash, D. P. 1976 "Male response to apparent female adultery in the mountain bluebird: an evolutionary interpretation". Am. Nat. 110, pp. 10971101.
Coon, C. S., Garn, S. M. & Birdsell, J. B. 1950 Races. Springfield, Ohio: C. Thomas.
Costa, R. & Bisol, P. M. 1978 "Genetic variability in deepsea organisms". Biol. Bull. 155, pp. 125133.
Darwin, C. 1872 The origin of species. London: John Murray.
Darwin, C. 1880 "Sir Wyville Thomson and natural selection". Nature, Lond. 23, p. 32.
Davitashvili, L. S. 1961 Teoriya polovogo otbora [Theory of sexual selection]. Moscow: Akademii Nauk.
Falconer, D. S. 1973 "Replicated selection for body weight in mice". Genet. Res. 22, pp. 291321.
Galton, F. 1909 Memories of my life. London: Methuen.
Gould, S. J. 1966 "Allometry and size in ontogeny and phylogeny". Biol. Rev. 41, pp. 587640.
Gould, S. J. 1971 "D'Arcy Thompson and the science of form". New Literary Hist. 2(2), pp. 229258.
Gould, S. J. 1974 "Allometry in primates, with emphasis on scaling and the evolution of the brain". In Approaches to primate paleobiology. Contrib. Primatol. 5, pp. 244292.
Gould, S. J. 1977 Ontogeny and philogeny. Cambridge, Mass.: Belknap Press.
Gould, S. J. 1978 "Sociobiology: the art of storytelling". New Scient. 80, pp. 530533.
Grassé, P. P. 1977 Evolution of living organisms. New York: Academic Press.
Gregory, W. K. 1936 "Habitus factors in the skeleton of fossil and recent mammals". Proc. Am. phil. Soc. 76, pp. 429444.
Harner, M. 1977 "The ecological basis for Aztec sacrifice". Am. Ethnologist. 4, pp. 117135.
Jerison, H. J. 1973 Evolution of the brain and intelligence. New York: Academic Press.
Lande, R. 1976 "Natural selection and random genetic drift in phenotypic evolution". Evolution 30, pp. 31334.
Lande, R. 1978 "Evolutionary mechanism of limb loss in tetrapods". Evolution 32, pp. 792.
Lewontin, R. C. 1978 "Adaptation". Scient. Am. 239 (3), pp. 156169. [Trad. it. in Storia naturale ed evoluzione, Le Scienze, Milano 1979, pp. 3949. Poi anche in Adattamento biologico, Le Scienze Quaderni, 27, 1985, pp. 313.]
Lewontin, R. C. 1979 "Sociobiology as an adaptationist program". Behav. Sci. 24, pp. 514.
Morton, E. S., Geitgey, M. S. & McGrath, S. 1978 "On bluebird 'responses to apparent female adultery'". Am. Nat. 112, pp. 968971.
Ortiz de Montellano, B. R. 1978 "Aztec cannibalism: an ecological necessity?". Science 200, pp. 611617.
Remane, A. 1971 Die Grundlagen des natürlichen Systems der vergleichenden Anatomie und der Phylogenetik. KönigsteinTaunus: Koeltz.
Rensch, B. 1959 Evolution above the species level. New York: Columbia University Press.
Riedl, R. 1975 Die Ordnung des Lebendigen. Hamburg: Paul Parey.
Riedl, R. 1977 "A systemsanalytical approach to macroevolutionary phenomena". Q. Rev. Biol. 52, pp. 351370.
Romanes, G. J. 1900 "The Darwinism of Darwin and of the postDarwinian schools". In Darwin, and after Darwin, vol. 2. London: Longmans, Green & Co.
Rudwick, M. J. S. 1964 "The function of zigzag deflections in the commissures of fossil brachiopods". Palaeontology 7, pp. 135171.
Sahlins, M. 1978 "Culture as protein and profit". New York review of books. 23 Nov., pp. 4553.
Schindewolf, O. H. 1950 Grundfragen der Paläontologie. Stuttgart: Schweizerbart.
Seilacher, A. 1970 "Arbeitskonzept zur Konstruktionsmorphologie". Lethaia 3, pp. 393396.
Seilacher, A. 1972 "Divaricate patterns in pelecypod shells". Lethaia 5, pp. 325343.
Shea, B. T. 1977 "Eskimo craniofacial morphology, cold stress and the maxillary sinus". Am. J. phys. Anthrop. 47, pp. 289300.
Stanley, S. M. 1970 "Relation of shell form to life habits in the Bivalvia (Mollusca)". Mem. Geol. Soc. Am. 125, 296 pp.
Sweeney, B. W. & Vannote, R. L. 1978 "Size variation and the distribution of hemimetabolous aquatic insects: two thermal equilibrium hypotheses". Science 200, pp. 444446.
Thompson, D. W. 1942 Growth and form. New York: Macmillan. [Trad. it. Crescita e forma, Boringhieri, Torino 1992.]
Waddington, C. H. & Cowe, J. R. 1969 "Computer simulation of a molluscan pigmentation pattern". J. theor. Biol. 25, pp. 219225.
Wallace, A. R. 1899 Darwinism. London: Macmillan.
Wilson, E. O. 1978 On human nature. Cambridge, Mass.: Harvard University Press. [Trad. it. Sulla natura umana, Zanichelli, Bologna 1980.]