L'adattamento è evidente anche nell'architettura generale dei principali gruppi di organismi. La struttura dell'ala degli uccelli o del corpo dei pesci ha meravigliato gli ingegneri, che hanno utilizzato queste creazioni naturali per dare forme ottimali alle macchine create dalla tecnologia (fig. 25.5).
L'adattamento è il punto di partenza di tutte le principali teorie dell'evoluzione; è l'aspetto principale della natura che ogni teoria soddisfacente deve spiegare. Essenzialmente la teoria darwiniana della selezione naturale propone che l'adattamento è originato da un cambiamento organico in risposta alla modificazione degli ambienti e non è un aspetto inalterabile dovuto all'immutabilità del mondo così come è stato creato:
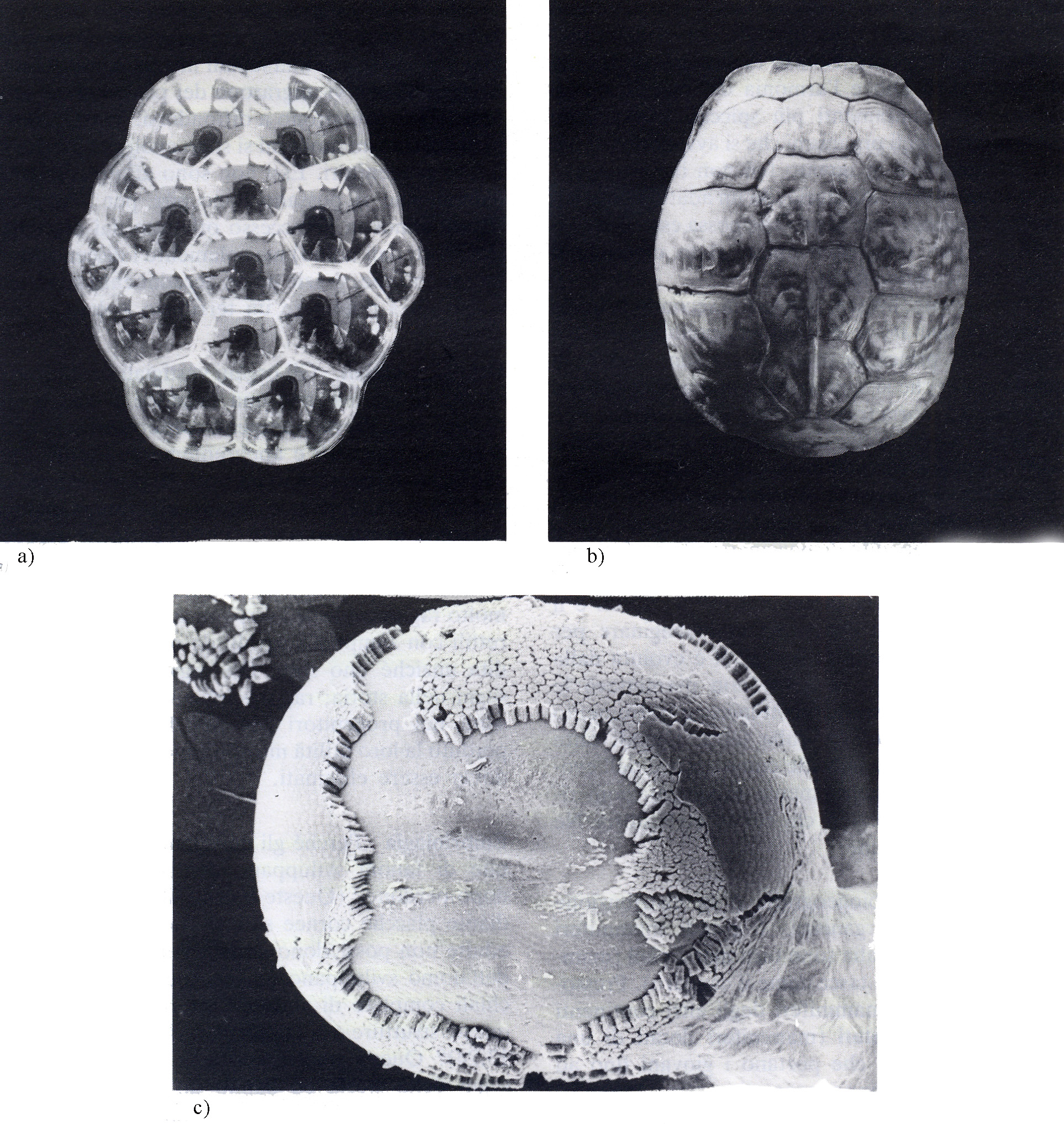
Questi stadi embrionali temporanei sono segni della storia ed hanno un senso solo se le balene con fanoni si sono evolute da balene con denti e se l'uomo si è evoluto dai pesci.
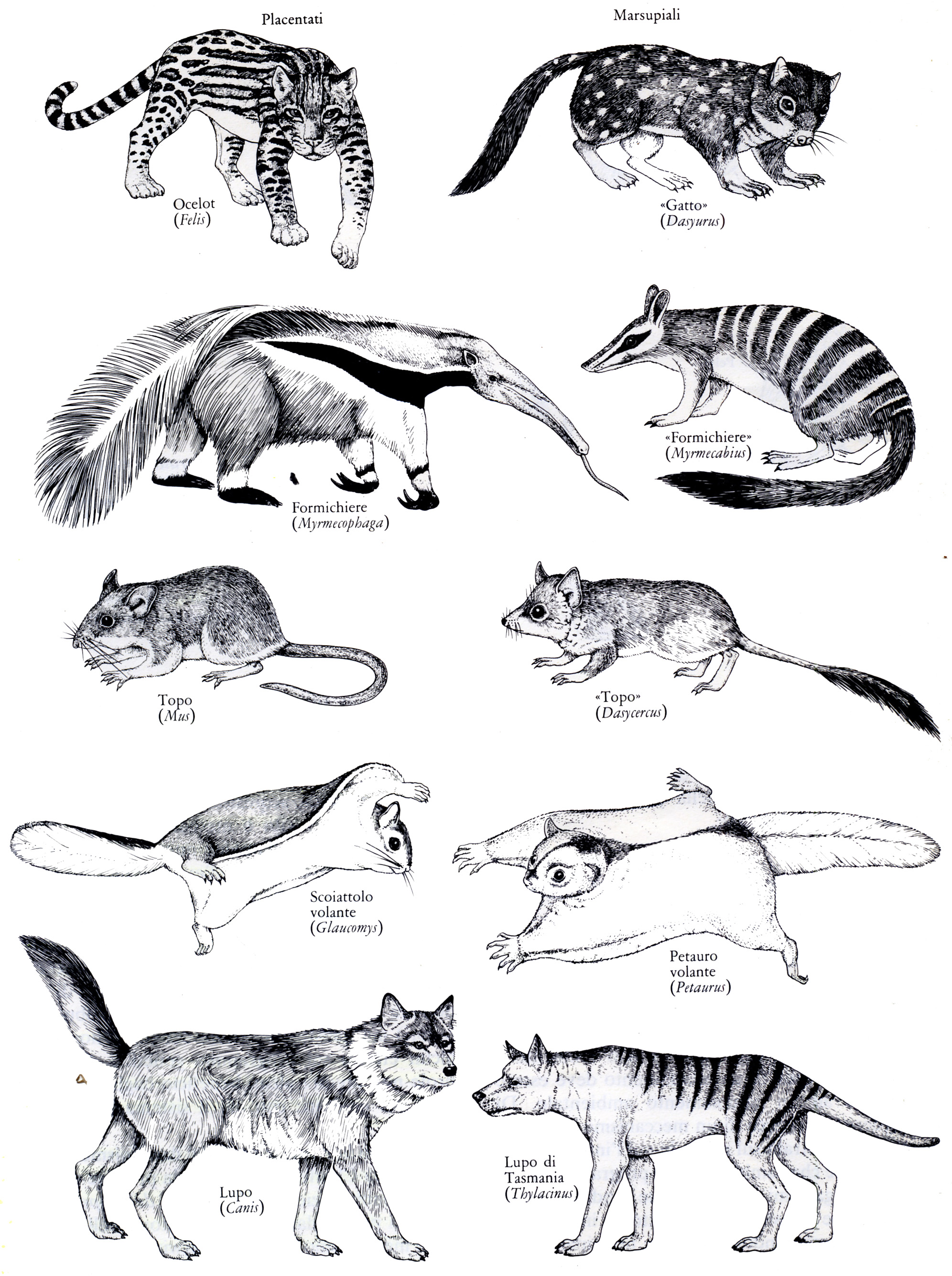
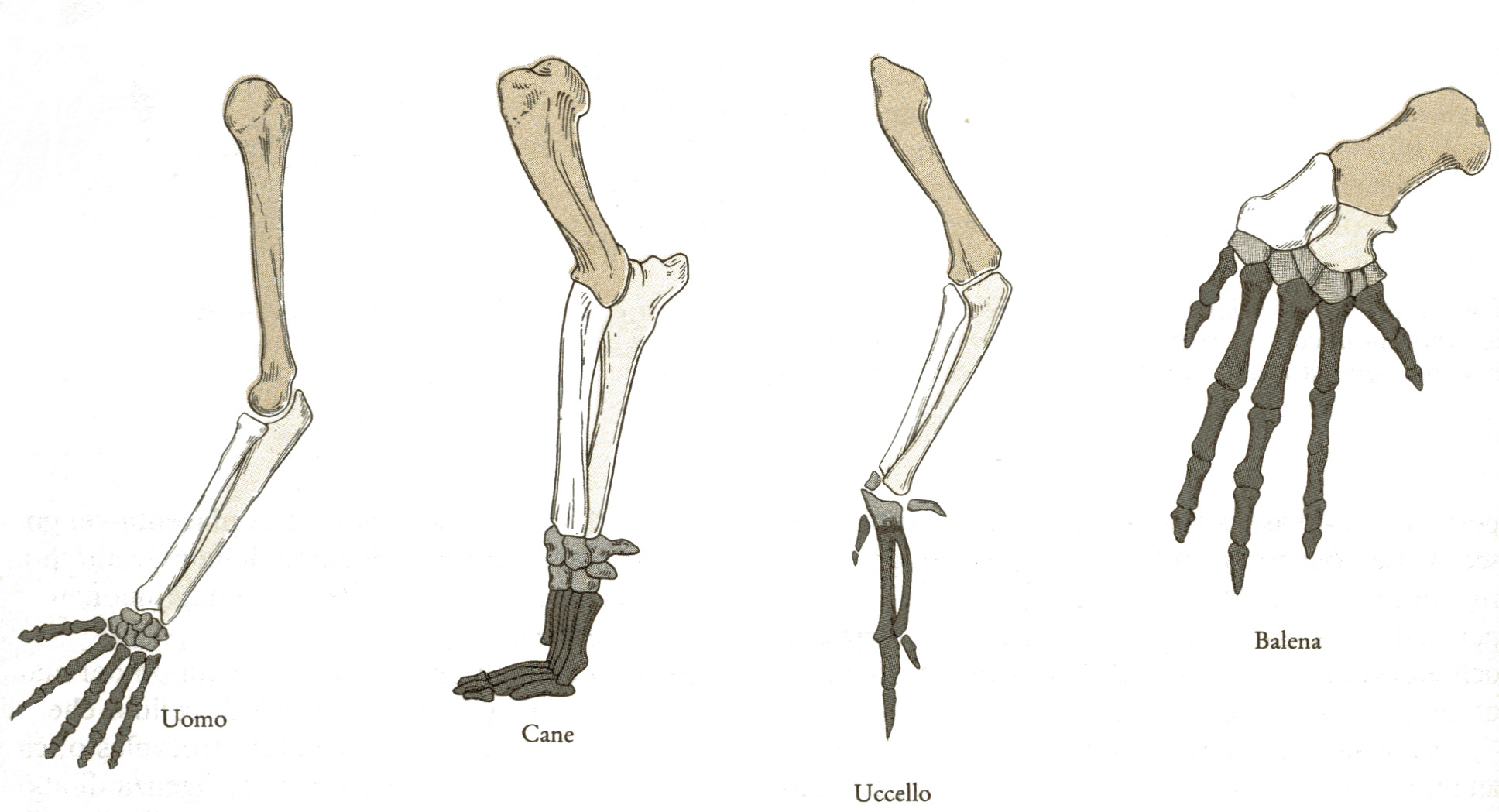
Perché Dio, al fine di costruire le migliori macchine per nuotare, volare o camminare, avrebbe utilizzato gli stessi elementi piegandoli e girandoli in modo così bizzarro? La struttura comune deve necessariamente riflettere la discendenza di tutti questi animali da un antenato che possedeva queste ossa.
L'evoluzione è provata dalle sue stesse imperfezioni. Quando, sulla base di queste, ammettiamo che essa è avvenuta, allora gli adattamenti più spiccati acquistano il valore di eccezionali testimonianze della finezza e della precisione con cui l'evoluzione può operare.
La prima di queste, la teoria lamarckista, è dovuta al biologo francese J.B. Lamarck che la pubblicò nel 1809, anno di nascita di Darwin. In questa teoria, che sostiene l'ereditarietà dei caratteri acquisiti, è previsto un processo semplice composto da una sola fase: gli organismi percepiscono le proprie necessità, acquisiscono gli adattamenti durante la propria vita e mediante i propri sforzi, trasmettono ereditariamente questi adattamenti alla propria discendenza. Se l'evoluzione operasse in questo modo sarebbe eccezionalmente efficiente e rapida; i sistemi genetici, però, non funzionano in questo modo. Nell'arco della propria vita un organismo può adattare il proprio corpo mediante l'esercizio (un esempio classico riportato nei libri di testo dell'era pre-industriale erano le braccia robuste dei fabbri) ma queste alterazioni del corpo non hanno alcun effetto sul programma genetico. Quindi la prole non eredita alcun beneficio dagli sforzi compiuti dai genitori: con ciò il sistema lamarckiano cade.
La seconda teoria, quella del darwinismo, prende il nome da Charles Darwin che la propose nel 1859 con la pubblicazione dell'Origine delle specie, uno dei libri più fondamentali della storia dell'occidente. Il meccanismo evolutivo darwiniano è più lento, più “sprecone” e meno efficiente di quello lamarckiano (fig. 25.8) ma ha in sé una concezione corretta del funzionamento dei programmi genetici.
Il sistema darwiniano dimostra che non sempre la natura segue il cammino più semplice che si possa immaginare, in quanto opera secondo un processo a due fasi in cui intervengono la variazione e la selezione naturale. Mediante la mutazione e la ricombinazione la variazione genetica fornisce il materiale grezzo; questa variazione è spontanea e casuale e, a differenza del sistema lamarckiano, non è incanalata nella direzione di un maggiore adattamento all'ambiente. La variazione, da sola, non produce evoluzione ma fornisce solamente il materiale grezzo per la seconda fase, quella della selezione naturale. Quando l'ambiente cambia si rendono vantaggiose nuove dimensioni, nuove forme, nuove funzioni e nuovi comportamenti. Gli organismi che variano in queste direzioni favorevoli tendono ad avere più successo nella riproduzione rispetto agli altri membri della specie; essi producono più prole e quindi, all'interno della specie, la frequenza dei geni per questi caratteri favorevoli aumenta. Questo processo, per cui i varianti favorevoli hanno un successo riproduttivo differenziale, è chiamato selezione naturale. Nel programma genetico non viene cambiato nulla direttamente dall'ambiente (come invece è nel lamarckiano); l'ambiente può solamente sottoporre a verifica, scegliere e conservare per selezione naturale. In questo senso il meccanismo darwiniano è un processo che riunisce il caso (la variazione) e la necessità (la selezione).
1. Tutti gli organismi di una specie sono diversi l'uno dall'altro (così come lo sono le persone che ci circondano).
2. Queste variazioni sono almeno in parte ereditate: i genitori alti tendono ad avere figli alti, i genitori neri figli, neri e i genitori bianchi figli bianchi.
3. Tutti gli organismi producono più prole di quella che può sopravvivere fino al momento della riproduzione. Darwin calcolò la rapidità di aumento degli animali con la riproduzione più veloce e più lenta nell'ipotesi che tutta la prole sopravvivesse fino alla riproduzione. Ad esempio una femmina di merluzzo può deporre diversi milioni di uova alla volta e sarebbero sufficienti pochi mesi perché i merluzzi riempissero gli oceani e trabordassero a terra. Anche per l'elefante, animale a riproduzione lenta, Darwin calcolò che se ogni esemplare vivesse cento anni e producesse sei figli durante la propria vita, in 750 anni una sola coppia genererebbe 19 milioni di discendenti.
4. Se gli organismi variano e non tutti possono sopravvivere, coloro che lo faranno saranno quelli che possiedono le variazioni ereditarie che aumentano il loro adattamento all'ambiente locale. Questo è il principio della selezione naturale ovvero della sopravvivenza del più adatto.
L'essenza della teoria di Darwin sta nell'affermazione che la selezione naturale non solo esiste effettivamente, ma è anche la forza creativa che dirige l'evoluzione. Per i creazionisti pre-darwiniani la selezione naturale era ammissibile quale meccanismo per la “purificazione del tipo”, cioè per l'eliminazione degli individui devianti che si scostavano eccessivamente dal tipo morfologico ideale ed immutabile. Per i darwinisti, viceversa, essa non è soltanto un esecutore bensì un artigiano creativo. La selezione naturale presiede al processo di modificazione organica accumulando le variazioni favorevoli poco a poco e generazione dopo generazione. Se ammettiamo il ruolo creativo della selezione naturale ci rendiamo conto che le quattro semplici affermazioni di Darwin da sole non coprono tutta la teoria; nell'ambito delle prime due, infatti, è necessario il verificarsi di altre quattro condizioni.
In primo luogo la variazione deve essere casuale e non indirizzata verso adattamenti favorevoli. Quando i biologi parlano di variazione “casuale” essi non intendono “ugualmente probabile in tutte le direzioni possibili” in quanto è ovvio che alcune mutazioni che si ripetono possono essere più frequenti di altre. Con il termine “casuale” si intende solamente che la variazione non si origina preferenzialmente in quelle direzioni in cui risulta essere vantaggiosa. Se ad un certo momento dimensioni minori diventano vantaggiose le mutazioni che provocano una diminuzione della statura non iniziano a verificarsi con maggiore frequenza ma mantengono il ritmo che avevano in precedenza; il loro aumento all'interno della popolazione è dovuto unicamente alla conservazione differenziale dovuta alla selezione naturale. Se le mutazioni si verificassero in modo preferenziale nelle direzioni vantaggiose, la selezione naturale sarebbe un semplice esecutore in quanto eliminerebbe i non adatti mentre gli adatti aumenterebbero di numero anche senza selezione: in questo caso la selezione sarebbe non-darwiniana. L'origine preferenziale delle mutazioni favorevoli è un'idea lamarckiana in quanto rappresenta l'espressione, in termini genetici, del pensiero di Lamarck (che, per quanto ne sappiamo, è sbagliato) secondo cui gli animali danno, alla percezione di determinate esigenze, risposte creative ed ereditabili.
In secondo luogo la variazione si deve verificare a tappe relativamente piccole. Infatti, se le specie si originassero all'improvviso, per il verificarsi fortuito di una grande mutazione che le rende immediatamente adatte al nuovo ambiente, la selezione ritornerebbe ad avere un ruolo esclusivamente esecutivo: quello di eliminare le specie parentali. La selezione non sarebbe una forza capace di dirigere in quanto gli adattamenti nascerebbero all'improvviso e senza il suo aiuto. Darwin insistette sul fatto che l'evoluzione si dovesse verificare a piccole tappe mediante incorporamenti successivi di picco cambiamenti per azione della selezione naturale l'adattamento viene costruito gradualmente grazi al potere amplificante della selezione naturale e n improvvisamente per eventi radicali e fortunosi.
Come terza condizione vi è che le popolazioni devono contenere una quota notevole di variabilità genetica. Se le variazioni ereditarie si produce raramente e la maggioranza dei membri di una popolazione fosse geneticamente uniforme, la velocità dell'evoluzione sarebbe limitata dalla variabilità anziché dalla selezione; in altre parole il cambiamento adattativo dovrebbe attendere che si producesse una quota di variabilità tale che la selezione vi possa agire. Normalmente secondo la teoria darwiniana una bassa velocità di evoluzione o una stabilità che si protrae per lunghi periodi sono l'effetto di una selezione debole o addirittura assente e mai di una carenza di variabilità.
Infine la teoria darwiniana ha ricevuto un grande sostegno dalla scoperta della natura discreta dell'ereditarietà, dovuta al fatto che i fattori ereditari sono particelle: i geni. Darwin credeva nell' “eredità per mescolamento”, cioè nell'idea che la prole dovesse mostrare caratteri intermedi rispetto a quelli parentali in quanto i fattori ereditari perdono la loro individualità al momento di fondersi nell'uovo fecondato.
L'eredità per mescolamento pose a Darwin un grande dilemma; secondo questa ipotesi un eventuale variante favorevole sarebbe all'inizio talmente raro che si dovrebbe accoppiare con un individuo della forma parentale. Nella prole della prima generazione la variazione risulterebbe ridotta della metà. Molto probabilmente anche questi individui si accoppierebbero con individui della forma parentale, e nella seconda generazione la variazione sarebbe ridotta ad un quarto. Alla fine la variazione favorevole verrebbe eliminata dal reincrocio con la forma parentale. La genetica mendeliana fornì la teoria dell'eredità discreta necessaria alla selezione naturale (fig. 25.10).
Un allele per un carattere vantaggioso non viene mai distrutto per reincrocio; se è recessivo esso può non essere espresso per alcune generazioni, ma non viene né diluito né distrutto. La selezione può provocare l'aumento della frequenza di un allele recessivo quando gli omozigoti manifestano il carattere vantaggioso.
Quindi, affinché la selezione naturale abbia il ruolo creativo che ha nella teoria darwiniana è necessario che la variazione genetica sia casuale, si realizzi a piccole tappe, sia frequente e sia discreta. Le mutazioni puntiformi della genetica mendeliana verificano queste condizioni in modo soddisfacente. Pertanto il moderno darwinismo viene indicato come teoria “sintetica” dell'evoluzione in quanto riunisce i postulati di Darwin sulla selezione naturale con la teoria genetica mendeliana, la quale soddisfa le condizioni sulla natura dell'ereditarietà.
Le conseguenze filosofiche della teoria darwiniana
459 Al momento della morte di Darwin, nel 1882, pressoché tutte le persone colte accettavano il fatto dell'evoluzione. Viceversa nello stesso periodo le opinioni di Darwin a proposito del meccanismo - cioè la teoria della selezione naturale - non avevano avuto altrettanto successo. Esse furono recepite da poche persone nel corso del diciannovesimo secolo e non riuscirono ad avere il sopravvento prima degli anni '30. Anche oggi la selezione naturale viene spesso male interpretata e trova resistenze fra i non-biologi. Perché questa idea, sostanzialmente semplice e potente, incontra simili difficoltà? La risposta è solo in parte di dominio della scienza empirica. Ad esempio abbiamo visto che la selezione naturale può essere spiegata in modo migliore da una teoria discreta dell'ereditarietà, e che questa fu disponibile solo dopo la riscoperta del lavoro di Mendel, nel 1900.
Ma un ostacolo ben più grande per l'accettazione della selezione naturale fu (ed è tuttora) il suo messaggio filosofico di fondo, capace di intaccare le idee tradizionali a proposito del progresso e della superiorità dell'uomo. La teoria darwiniana della selezione naturale ha turbato molte persone e ha provocato ilarità in altre perché essa insiste sul fatto che il cammino dell'evoluzione e dell'armonia della natura “non ha un fine”. In precedenza tutto sembrava avere un ruolo prefissato nell'armonia della natura. In una colonia di formiche le operaie raccolgono il cibo, i soldati provvedono alla difesa e la regina alla procreazione; le api si cibano di nettare e a loro volta impollinano i fiori. Questa armonia del vantaggio reciproco può riflettere qualcosa che non sia un principio “superiore” che agisce a livello dell'ecosistema, della colonia di formiche o di tutta la natura? Darwin negò vigorosamente l'azione di qualsiasi principio “superiore” nella natura. La selezione naturale non è altro che la lotta degli individui per sopravvivere e per perpetuare i propri geni nelle generazioni future. Ogni parvenza di un ordine superiore si origina come sottoprodotto da questa lotta fra individui. Né la selezione, né tanto meno l'ecosistema sono a conoscenza del “bene della specie”. Le api ed i fiori sono adattati per il proprio vantaggio e non per il vantaggio altrui o per un dato ordine naturale - le api mangiano e i fiori vengono impollinati. Il sistema “cooperativo” rappresenta solamente un punto di contatto fra gli individui di due specie che lottano esclusivamente per se stessi e non collaborano per alcuna armonia reciproca.
Il secondo punto fondamentale è che Darwin intuì che nell'evoluzione non è insito il progresso: la vita non si muove affatto inesorabilmente da stati “inferiori” a stati “superiori”; non vi è alcuna scala della vita o catena dell'essere. Nessuna delle cattive interpretazioni dell'evoluzione è più diffusa di quella secondo cui l'evoluzione porta ad un progressivo miglioramento della vita. La maggioranza delle teorie evolutive non darwiniane postula un progresso immanente. Lamarck, ad esempio, sosteneva che gli organismi rispondono creativamente alle loro necessità tramite “una forza che tende incessantemente a complicare l'organizzazione”. Invece la selezione naturale può solo adattare gli organismi agli ambienti locali; il suo solo “progresso” è una migliore sintonia fra l'organismo e le condizioni locali - ad esempio il portare all'evoluzione di un elefante con lunghi peli (il mammuth) che possa vivere nei climi glaciali. La selezione naturale ha portato ad un aumento delle dimensioni del cervello in diverse linee evolutive dei mammiferi, mentre in molti parassiti ha prodotto una degenerazione morfologica estrema - ad esempio gli adulti di Sacculina sono ridotti a contenitori di organi riproduttivi mentre gli stadi larvali non parassiti mostrano le complessità anatomiche proprie del loro gruppo (fig. 25.11).

Chi può dire quale dei due adattamenti sia il “migliore”? Sia i mammiferi con grande cervello che i parassiti degenerati sono ben adattati e, nel loro ambiente, hanno pieno successo. Darwin scrisse: “Dopo lunga riflessione non posso abbandonare la convinzione che non esiste alcuna tendenza verso uno sviluppo progressivo”.
Un terzo aspetto, ancora più in contrasto con le idee convenzionali del tempo, fu che Darwin sostenne con convinzione il materialismo filosofico, cioè quella concezione secondo cui la materia è alla base di tutta l'esistenza e che lo “spirito” e la “mente” sono prodotti o invenzioni di un cervello materiale. Darwin sostenne una spiegazione completamente naturalistica della vita ricusando così una delle tradizioni più radicate nel pensiero occidentale: la separazione fra mente e materia e l'attribuzione di uno stato superiore alla prima. Egli concepì la mente umana come il risultato naturale delle pressioni selettive che favorivano un cervello grande e complesso. Egli non attribuì all'uomo alcuna collocazione privilegiata in quanto lo concepì inestricabilmente connesso con il resto della vita. Darwin, con la sua spiegazione, non voleva demolire le convinzioni religiose di nessuno; egli volle solamente affermare il fatto che le cause e gli attributi divini non hanno niente a che fare con l'indagine scientifica. La sua visione del cervello umano era quella di un oggetto di studio scientifico, come qualsiasi altra parte della natura.
In un suo famoso brano Sigmund Freud sottolineò che il risultato principale dell'avanzamento della scienza è la diminuzione di quello stato di esaltazione che abbiamo edificato a nostro beneficio. Inizialmente l'astronomia e la fisica ci dimostrarono che noi non abitiamo un corpo situato al centro dell'universo bensì un piccolo pianeta che ruota attorno ad una stella insignificante alla periferia di una galassia fra i molti milioni che ne esistono. Successivamente la biologia dimostrò che non siamo stati creati a somiglianza di un Dio onnipotente ma ci siamo evoluti dalle scimmie grazie allo stesso processo che regola la storia di tutti gli organismi. Infine, la psicologia starebbe a dimostrare che le nostre menti non sono macchine razionali ma miscugli immensamente complessi di impulsi, sentimenti, desideri repressi ed apprendimenti.
Nessun uomo più di Darwin ha contribuito a questa ritirata dalla nostra arroganza cosmica. Affermando che noi non siamo altro che un prodotto di un processo naturale che non ha né scopo né direzione, Darwin ci ha costretti a cercare un significato all'interno di noi stessi e non nella natura che ci circonda. Mettendo in risalto la nostra unitarietà con tutto il mondo della vita egli ha incoraggiato un'umiltà che noi, con la nostra tecnologia minacciosa, faremmo bene ad imparare.
L'evoluzione dell'altruismo: un banco di prova per la concezione darwiniana
460 L'insistenza di Darwin sui vantaggi dell'individuo pone degli interessanti problemi agli studiosi del comportamento sociale. Le azioni altruistiche, cioè quei comportamenti che abbassano il successo riproduttivo del singolo individuo a favore di un apparente vantaggio degli altri, sono il cemento delle società stabili. L'ape operaia rinuncia alla riproduzione per aiutare la regina, cioè sua madre, ad allevare più sorelle. Fra le ghiandaie della Florida spesso i giovani maschi si prendono cura dei nidi altrui anziché allevare i propri piccoli. Come si possono evolvere simili comportamenti se la selezione opera unicamente per un aumento del successo riproduttivo dei singoli individui?
La selezione di gruppo
461 Una possibile risposta, in apparente contrasto con il postulato darwiniano della selezione individuale, sostiene che la selezione spesso opera su interi gruppi intesi come unità. Si supponga che una specie sia divisa in 100 sottogruppi e che il corso futuro della sua evoluzione dipenda dalla sopravvivenza differenziale dei sottogruppi anziché degli individui. In questo caso un gene che porta al sacrificio di un individuo per il bene del sottogruppo verrebbe mantenuto grazie alla selezione di gruppo: i gruppi che non contengono individui portatori del gene altruista verrebbero sostituiti dai gruppi con individui votati al sacrificio. Un esempio classico è costituito dalle grida d'allarme in alcuni uccelli gregari. In alcune specie che formano grandi stormi, singoli individui, alla vista di un predatore, emettono il grido di allarme che provoca la frammentazione del gruppo. Sembrerebbe quindi che questi individui si mettano a repentaglio a vantaggio dei loro compagni di gruppo.
A molti la selezione di gruppo potrebbe apparire corretta, ma essa non ha avuto molto seguito per due ragioni fondamentali. Innanzitutto poche sono le specie che sono suddivise in gruppi sufficientemente numerosi e sufficientemente isolati per comportarsi come vere e proprie unità nei confronti della selezione. In secondo luogo le “storie” evolutive che vengono spiegate in base alla selezione di gruppo possono essere spiegate anche in termini di selezione individuale. Siamo sicuri che l'emissione di un grido d'allarme non rappresenti un vantaggio anche per colui che lo emette oltreché per i suoi compagni di gruppo? A questo proposito si possono supporre molti vantaggi per l'individuo che dà l'allarme: il movimento del gruppo provocato dall'allarme potrebbe confondere il predatore rendendo così meno probabile la cattura di ogni individuo compreso quello che ha dato l'allarme (si può anche immaginare che il ruolo di individuo allarmante sia peggiore di quello di individuo silenzioso ma, se nessuno desse l'allarme, il predatore catturerebbe sempre qualcuno nel gruppo riducendo così la sopravvivenza individuale). Un'altra funzione dell'allarme potrebbe essere quella di far spostare il gruppo verso una posizione più sicura. In questo caso si può obiettare che per l'uccello che dà l'allarme potrebbe essere meglio mettersi al sicuro da solo e far sopportare l'attacco al resto del gruppo. Tuttavia, spesso i predatori vengono attratti dagli individui che si trovano fuori dai ranghi così come, durante una sfilata, noi veniamo colpiti da quelli che non tengono il passo; quindi, nel muoversi verso un luogo più sicuro, è meglio se lo si fa tutti insieme. Tutte queste considerazioni non vogliono sostenere la superiorità delle spiegazioni date in base alla selezione individuale rispetto a quelle date in base alla selezione di gruppo, ma solamente mostrare che quei casi di sacrificio apparente possono essere interpretati normalmente anche in termini di vantaggio individuale darwiniano.
La kin selection (selezione familiare)
462 Ci troviamo di fronte a molti altri casi di altruismo che sfuggono alla spiegazione in termini di successo riproduttivo del singolo individuo. Non v'è dubbio che spesso alcuni individui rinunciano alla propria riproduzione per aiutare altri ad allevare la prole: l'ape operaia, essendo sterile, non può trasmettere i propri geni ad alcun discendente. Come si accorda questo comportamento con la necessità darwiniana del vantaggio individuale? Negli anni '60 ii biologo inglese W.D. Hamilton ha offerto una soluzione elegante per questo serio problema. Egli considerò che il vantaggio individuale (o fitness) dovesse essere misurato dal successo ottenuto nel trasmettere i geni alle generazioni future. Un individuo non può trasmettere tutti i propri geni in quanto, nella risproduzione sessuata, i geni della prole sono per metà materni e per metà paterni. Tuttavia nei parenti di un individuo sono presenti copie dei suoi geni e più stretta è la parentela maggiore è la percentuale di geni in comune.
Fra gli organismi diploidi che si riproducono sessualmente i fratelli hanno metà dei loro geni in comune (se uno ha il gene materno A anche sua sorella avrà il 50% di probabilità di averlo perché l'uovo aploide da cui essa si è sviluppata poteva contenere o lo stesso cromosoma con A o l'altro cromosoma di quella coppia; estendendo il ragionamento a tutti i geni si ottiene il valore medio del 50%). Quindi se un individuo si viene a trovare in una situazione in cui può salvare se stesso o morire per salvare tre fratelli, per la propria fitness egli dovrà sacrificarsi. In questo modo, grazie al fatto che i suoi fratelli potranno continuare a riprodursi, egli trasmetterà il 150% dei propri geni. In altre parole le azioni “altruistiche” eseguite a beneficio di un parente sufficientemente prossimo sono in realtà atti egoistici a vantaggio dei propri geni: questa idea è chiamata kin selection (o selezione familiare).
Possiamo controllare la teoria della kin selection (una soluzione darwiniana al problema dell'altruismo) esaminando se in natura gli apparenti atti di altruismo vengono eseguiti a vantaggio di parenti sufficientemente prossimi. Ad esempio sappiamo che fra le ghiandaie della Florida quei nidi che vengono curati dai genitori e da aiutanti aggiuntivi producono più giovani di quelli che contengono lo stesso numero di uova ma sono curati dai soli genitori. Dal momento che gli aiutanti sono fratelli e sorelle dei genitori, essi (gli aiutanti) aumentano probabilmente la propria fitness incrementando il successo di copie dei loro geni nella prole dei propri fratelli.
La kin selection ha dato anche un'interessante spiegazione al mistero della presenza di operaie sterili fra gli insetti dell'ordine Hymenoptera, che comprende le formiche, le api e le vespe. In queste specie sociali le operaie sono geneticamente femmine, ma sono incapaci di produrre prole. Esse passano la loro vita aiutando la madre (la regina) ad allevare più sorelle, curando e nutrendo le uova e le larve e sorvegliando e difendendo il nido (le operaie più grandi, dette “soldati”, costituiscono un “esercito” di femmine). Perché le caste sterili sono sempre formate da femmine? Perché questo comportamento sociale si è evoluto per non meno di 11 volte indipendentemente fra gli imenotteri e solo un'altra volta (nelle termiti) in tutto il resto degli insetti? La risposta potrebbe risiedere nel sistema genetico atipico degli imenotteri.
Negli imenotteri le femmine si sviluppano da uova fecondate e sono diploidi come nell'uomo, mentre i maschi si sviluppano da uova non fecondate; i maschi, quindi, sono aploidi e contengono soltanto un corredo di geni materni: questo sistema genetico è indicato con il termine di aplodiploidia. Consideriamo ora la parentela genetica che una formica femmina ha con le proprie sorelle e con la madre (fig. 25.12).
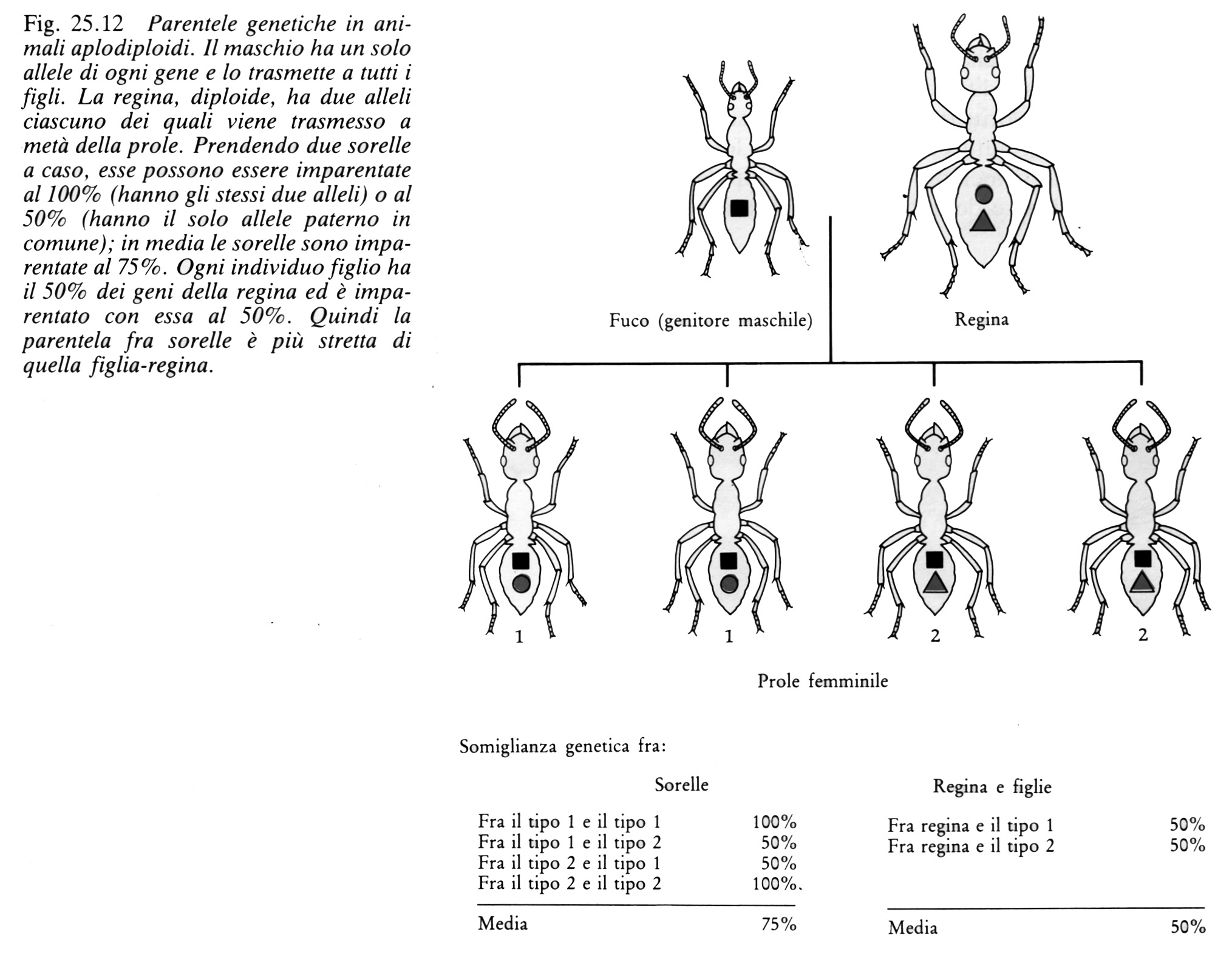
Tutta la prole di una regina ha metà dei geni di questa e quindi ogni operaia è imparentata con sua madre al 50%. E per quanto riguarda la parentela fra operaie? Consideriamo il gene A. Qual è la probabilità che due sorelle lo abbiano in comune? Se A è un gene materno ogni sorella ha una probabilità del 50% di riceverlo, come nelle specie diploidi. Se, invece, è un gene paterno ogni sorella lo deve avere poiché tutti i padri sono aploidi e trasmettono tutti i propri geni a tutte le figlie. Quindi la parentela fra due sorelle è in media del 50% per i geni materni e del 100% per quelli paterni cioè, complessivamente, del 75%. Nelle specie aplodiploidi le femmine sono più strettamente imparentate con le proprie sorelle di quanto sarebbero con la propria prole potenziale! Quindi per loro potrebbe risultare vantaggioso rinunciare alla riproduzione per aiutare la madre, cioè la regina, ad allevare più sorelle (queste argomentazioni sono valide solo se tutte le operaie del nido hanno lo stesso padre cioè solo se la regina viene fecondata una volta sola ed accumula lo sperma per produrre tutte le figlie; non sembra che fra gli imenotteri sia presente l'inseminazione singola e i molti casi noti di inseminazione multipla - in cui lo sperma proviene da più padri - mettono in difficoltà questa teoria). Le caste sterili sono formate da femmine perché i maschi non hanno questa possibilità di una maggior parentela con i fratelli (datene una spiegazione). Questa forma di società è favorita solo nelle specie aplodiploidi in quanto in quelle diploidi ordinarie (come l'uomo) non c'è questa asimmetria di parentele. Noi siamo ugualmente imparentati con i nostri fratelli e con la nostra prole.
Altri agenti del cambiamento evolutivo
463 Il risultato della selezione naturale è un successo riproduttivo differenziale. In termini genetici gli organismi favoriti dalla selezione trasmettono, alle generazioni future, più copie dei loro geni rispetto agli altri membri della stessa popolazione (una popolazione è definita come qualsiasi gruppo discreto in cui gli individui si incrociano liberamente,). La somma di tutti i geni presenti in tutti i membri di una popolazione è detta pool genetico. Se la selezione favorisce ad esempio gli organismi che portano i geni per dimensioni minori, il numero relativo (cioè la frequenza) di questi geni all'interno del pool aumenterà. Così i genetisti definiscono spesso l'evoluzione come una modificazione della frequenza genica all'interno dei pool genetici. Di conseguenza potremmo specificare le cause dei cambiamenti evolutivi all'interno delle popolazioni elencando tutti quegli agenti che provocano modificazioni nelle frequenze geniche (in questa trattazione escludiamo quelle modificazioni locali delle frequenze geniche provocate dall'emigrazione o dall'immigrazione di individui da altre popolazioni della stessa specie; questi spostamenti alterano la distribuzione delle frequenze geniche nelle popolazioni locali, ma non modificano la frequenza genica complessiva della specie).
Alcuni biologi affermano che la selezione naturale è il solo agente importante che possa portare ad un cambiamento sostanziale delle frequenze geniche nelle popolazioni. Siamo attorniati da troppi esempi di architettura meravigliosa nel mondo organico per negare l'enorme influenza di controllo esercitata dalla selezione naturale, soprattutto considerando che non si conoscono altri meccanismi di cambiamento adattativo. A questo proposito rimaniamo impressionati dagli organi elaborati per le esibizioni e i combattimenti ritualizzati presenti in molti animali (fig. 25.13).

Questi organi non migliorano la struttura meccanica dell'animale e non gli sono di aiuto né per nutrirsi né per muoversi; anzi, potrebbero risultare svantaggiosi in quanto attingono alle risorse metaboliche dell'organismo (le corna di un alce estinto dell'Irlanda pesavano fino a 36 kg ed erano impiantate su un cranio di soli 2,2 kg; esse venivano perdute e riformate ogni anno). Questi organi sono però favoriti dalla selezione naturale per la ragione più diretta: essi aumentano il successo riproduttivo consentendo un maggior numero di rapporti sessuali con un maggior numero di partner. I maschi con le corna più grandi, con le pinne più vistose o con il canto più elaborato, vengono scelti dalle femmine o possono scacciare più efficacemente gli altri maschi. Se l'evoluzione non fosse controllata dalla selezione naturale e non si misurasse in termini di successo riproduttivo, sarebbe difficile spiegare queste strutture così elaborate e ingombranti.
Malgrado tutto ciò negli ultimi anni i progressi della genetica molecolare hanno fatto nascere dubbi sull'idea che l'unico agente importante del cambiamento evolutivo sia la selezione naturale. I cambiamenti prodotti dalla selezione naturale sono deterministici e adattativi. Oggi molti biologi ritengono che vi siano altri cambiamenti evolutivi importanti e che questi siano casuali e neutrali. Il darwinismo classico ammette, in qualche rara situazione, cambiamenti casuali, ma forse questi avvengono assai più di frequente e in un più ampio spettro di circostanze.
Prima di proseguire dobbiamo fare una distinzione fra variazione casuale e cambiamento casuale. La variazione casuale è il materiale grezzo su cui la selezione naturale agisce per produrre il cambiamento evolutivo; il cambiamento casuale è quella modificazione direzionale delle frequenze geniche che si produce in assenza di selezione naturale per le stesse ragioni per cui circa una volta su 1024 ci capita di ottenere 10 teste o 10 croci consecutivamente con il lancio di una moneta.
Il principio del fondatore
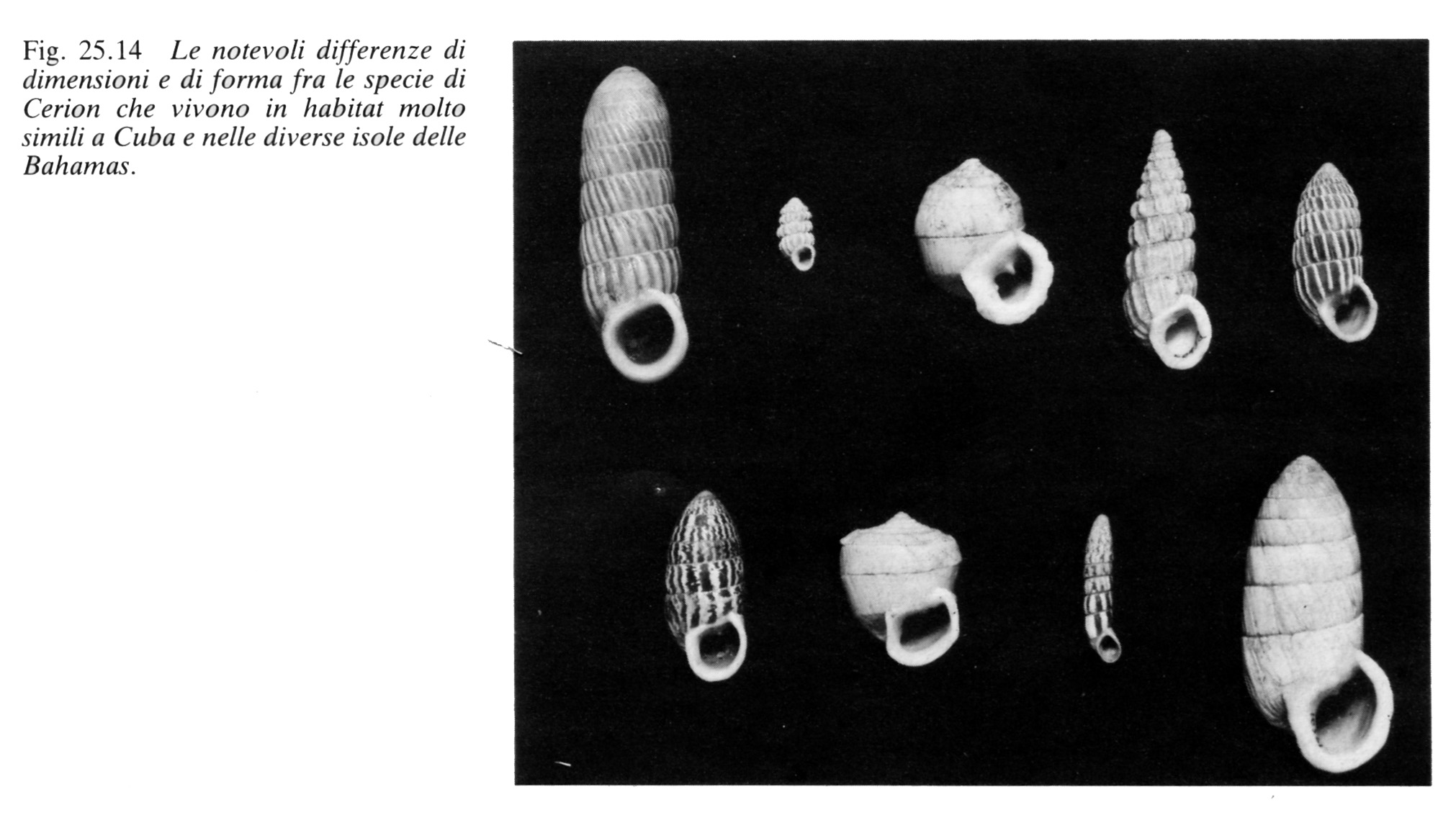
464 In tutte le 700 isole dell'arcipelago delle Bahamas vivono chiocciole terrestri del genere Cerion (fig. 24.14).
Durante le epoche glaciali, quando il livello del mare era più basso, molte di queste isole erano in comunicazione e le chiocciole poterono migrare da un luogo all'altro. Molte altre isole, invece, sono circondate da mari profondi e quindi le chiocciole possono esservi arrivate solamente con sistemi accidentali e occasionali: su tronchi galleggianti, a causa di uragani, o nel fango che rimane attaccato alle zampe degli uccelli marini (succede anche questo). Poiché queste chiocciole sono ermafroditi potenzialmente capaci di effettuare l'autofecondazione, singoli individui possono fondare delle colonie.
Supponiamo ora che tempo addietro in una popolazione parentale sia insorto, per una rara mutazione, un gene favorevole per la colorazione scura e che, per effetto della selezione naturale, oggi sia presente in metà delle chiocciole di una data isola. Supponiamo anche che un uragano abbia portato due chiocciole all'isola più vicina dove esse si sono moltiplicate ed hanno dato luogo ad una nuova popolazione. Gli ambienti delle due isole sono simili ed in entrambe il gene per la colorazione scura è favorito. L'uragano, però, “ha raccolto” le due chiocciole a caso. Dal momento che il gene favorevole era presente solo nella metà delle chiocciole, vi è una probabilità del 25% che nessuna delle due chiocciole colonizzatrici lo abbia portato nella nuova isola, anche se qui esso sarebbe favorito (si ottiene il 25% cioè 0,25 moltiplicando 0,5 per 0,5: questa è infatti la probabilità che ciascuna chiocciola non porti il gene favorevole; per lo stesso motivo la probabilità di ottenere due croci consecutivamente con una moneta è del 25%). Dal momento che bisogna considerare migliaia di geni è inevitabile che per alcuni di essi le piccole popolazioni fondatrici abbiano frequenze alleliche molto diverse da quelle delle popolazioni parentali: ciò è un semplice effetto del caso; inoltre è pressoché inevitabile che vengano perduti geni vantaggiosi. Questo effetto è detto principio del fondatore. Secondo questo principio quando le popolazioni sono sufficientemente piccole intervengono eventi puramente casuali che alterano le frequenze geniche. Per usare l'esempio della moneta, immaginiamo una grande popolazione parentale costituita da 100 000 lanci: la frequenza delle teste e delle croci sarà vicina al 50%. Se noi estraiamo una nuova popolazione di 5 000 lanci possiamo essere sicuri che la frequenza delle teste e delle croci sarà simile a quella della popolazione parentale, ma se estraiamo due soli lanci come “colonizzatori” avremo una probabilità del 50% di ottenere scostamenti notevoli nella proporzione fra teste e croci (25% di ottenere due teste e 25% di ottenere due croci).
Quasi tutte le 700 isole delle Bahamas ospitano forme differenti di Cerion e possiamo essere pressoché certi che queste differenze non si sono originate per selezione naturale.
La deriva genetica
465 Molte popolazioni subiscono grandi fluttuazioni numeriche e sono sottoposte a periodi più o meno regolari di espansione e diminuzione. Durante questi periodi di diminuzione la popolazione può diventare molto piccola e questa situazione viene indicata con la espressione collo.di-bottiglia (bottle-neck).
Consideriamo ora il destino potenziale di un gene favorevole che produce antenne lunghe in una popolazione di 12 scarabei sottoposta ad un collo di bottiglia. Supponiamo che 3 di essi abbiano il gene e che questo conferisca un vantaggio selettivo del 25% (cioè lo scarabeo con il gene abbia un 25% di prole in più rispetto allo scarabeo senza il gene). Sfortunatamente un elefante che passa di lì schiaccia metà della popolazione (una piccola differenza nella lunghezza delle antenne non protegge da calamità di questo tipo). Ora, la probabilità che il gene sia stato eliminato perché fra i 6 scarabei morti sono compresi i 3 che lo portavano, è grande.
In una popolazione diffusa, composta da 12 milioni di scarabei, un eventuale gene favorevole presente in un quarto degli individui quasi certamente si diffonderà fino a prevalere, mentre nelle popolazioni piccole esso può essere perduto semplicemente a causa della morte accidentale. Queste fluttuazioni casuali delle frequenze geniche prendono il nome di deriva genetica (in questo esempio l'elefante rappresenta una causa di morte casuale, almeno in rapporto all'effetto del gene per le antenne).
Le piccole popolazioni di colonizzatori sono particolarmente soggette alle alterazioni delle frequenze geniche dovute a processi casuali che non sono influenzati dalla selezione naturale (ma che spesso si oppongono ad essa). In primo luogo, per il principio del fondatore, i colonizzatori portano un campione incompleto del pool genetico parentale e inoltre, per effetto della deriva genetica, subiscono fluttuazioni casuali delle frequenze geniche. Non ci dobbiamo sorprendere, quindi, del fatto che le nuove specie nascano così spesso sulle isole.
La scuola rigidamente selezionista del darwinismo classico ammetteva il principio del fondatore e la deriva genetica, ma solo come fattori di importanza secondaria in quanto, apparentemente, essi possono agire solo su popolazioni molto piccole. Tuttavia recentemente vi sono state nuove acquisizioni nelle teorie evoluzionistiche che hanno portato ad un ridimensionamento di questa fiducia incondizionata nella selezione. Innanzitutto il ruolo delle piccole popolazioni nell'origine delle specie può avere un'importanza maggiore di quella attribuitagli nelle teorie precedenti. In secondo luogo studi recenti sulla struttura geografica delle specie hanno dimostrato che molte di queste (se non addirittura la maggioranza) sono suddivise in piccole sottopopolazioni con scarsi contatti da migrazione. Queste sottopopolazioni hanno spesso dimensioni tali da rendere efficace la deriva genetica e si possono comportare come vere e proprie unità evolutive.
Immaginiamo una specie di farfalle che viva solo alle alte quote; se le montagne sono sufficientemente distanziate in modo che gli individui non si possano spostare da una all'altra, e se su ciascuna di esse vivono solo poche farfalle, allora il fatto che la specie sia formata complessivamente da un milione di individui può non avere alcuna importanza: le poche farfalle di ogni montagna si comporteranno come un'unità evolutiva indipendente. In terzo luogo sono state proposte nuove ipotesi di cambiamenti non selettivi delle frequenze geniche fondate sull'idea della pressione di mutazione.
La pressione di mutazione e la teoria del neutralismo
466 Il codice genetico è degenerato rispetto alla terza posizione di molte triplette. Ad esempio nell'RNA messaggero le triplette GUU, GUC, GUA e GUG specificano tutte l'aminoacido valina. La selezione può non agire su una mutazione che porta da GUC a GUA, in quanto essa “guarda” unicamente all'organismo finito e non ai suoi geni. Se la valina può essere prodotta indifferentemente da ciascuna di queste quattro triplette, la mutazione che porta da una all'altra può risultare invisibile ai processi darwiniani: questo cambiamento è considerato una mutazione neutrale. Esistono due modi con i quali una simile mutazione si può diffondere all'interno della popolazione: la deriva genetica e la pressione di mutazione.
La deriva genetica opera più facilmente nelle popolazioni piccole. Nelle popolazioni grandi è raro che un gene sfavorevole abbia il sopravvento, poiché la probabilità di una morte casuale di tutti gli individui avvantaggiati è vicina a zero. Viceversa, se una mutazione è realmente neutrale essa non sarà suscettibile all'azione della selezione e rimarrà nella popolazione con una frequenza determinata dai tassi di formazione e di scomparsa. Le fluttuazioni di questa frequenza sono dovute unicamente alla casualità della deriva genetica. In milioni di anni questa mutazione può avere il sopravvento anche se la popolazione conserva le sue grandi dimensioni (purché vi sia tempo a sufficienza con una moneta si può ottenere una serie di 100 croci).
Supponiamo ora che i membri di una nuova specie incorporino valina mediante la tripletta GUA e che le mutazioni da GUA a GUU siano più frequenti di quelle da GUA a GUC o GUG mentre la retromutazione da GUU ad una delle altre tre triplette abbia una frequenza relativamente bassa. Dal momento che la selezione non può interferire in questo processo, alla fine GUU raggiungerà un'alta frequenza nel pool genetico semplicemente a causa della pressione di mutazione. Poiché la teoria darwiniana sostiene che la mutazione è solo il materiale grezzo e l'agente del cambiamento delle frequenze geniche è la selezione, questo ipotetico aumento di GUU dovuto alla pressione di mutazione sarebbe un processo non-darwiniano.
Cosa accade delle mutazioni leggermente nocive che vengono lentamente rimosse dalla selezione ma si formano più rapidamente per effetto del ripetersi degli eventi mutativi? Queste mutazioni rimarranno nelle popolazioni malgrado il loro svantaggio selettivo e, come quelle realmente neutrali, potranno prevalere per effetto della deriva genetica. In breve, affinché un gene sia mantenuto in opposizione alla influenza attiva della selezione che tende a rimuoverlo, è sufficiente che la pressione di mutazione sia maggiore della pressione di selezione.
Indicazioni della neutralità selettiva di buona parte della variabilità genetica
467 Per sostenere che buona parte del cambiamento evolutivo non è sotto il controllo della selezione sono state sfruttate due categorie di prove: l'orologio evolutivo e la quota di variabilità nelle popolazioni naturali.
L'orologio evolutivo
Il decadimento radioattivo può definire l'età delle rocce perché è un processo casuale. Tutti gli atomi dell'isotopo radioattivo potassio-40 hanno la stessa probabilità di decadere ad argon-40 entro un certo intervallo di tempo. Quindi, dopo un certo tempo, metà degli atomi di partenza saranno scomparsi e metà di quelli rimanenti scompariranno nel successivo intervallo di semivita. Il processo continua fino a che tutti gli atomi radioattivi saranno scomparsi. Se conosciamo la velocità di decadimento possiamo calcolare l'età di una roccia misurando il rapporto fra gli atomi residui e quelli originali (cioè quelli residui più quelli formatisi per decadimento).
Se l'evoluzione molecolare procedesse ad una velocità costante potremmo datare la storia della vita così come datiamo le rocce in base alla velocità costante del decadimento radioattivo (fig. 25.15).
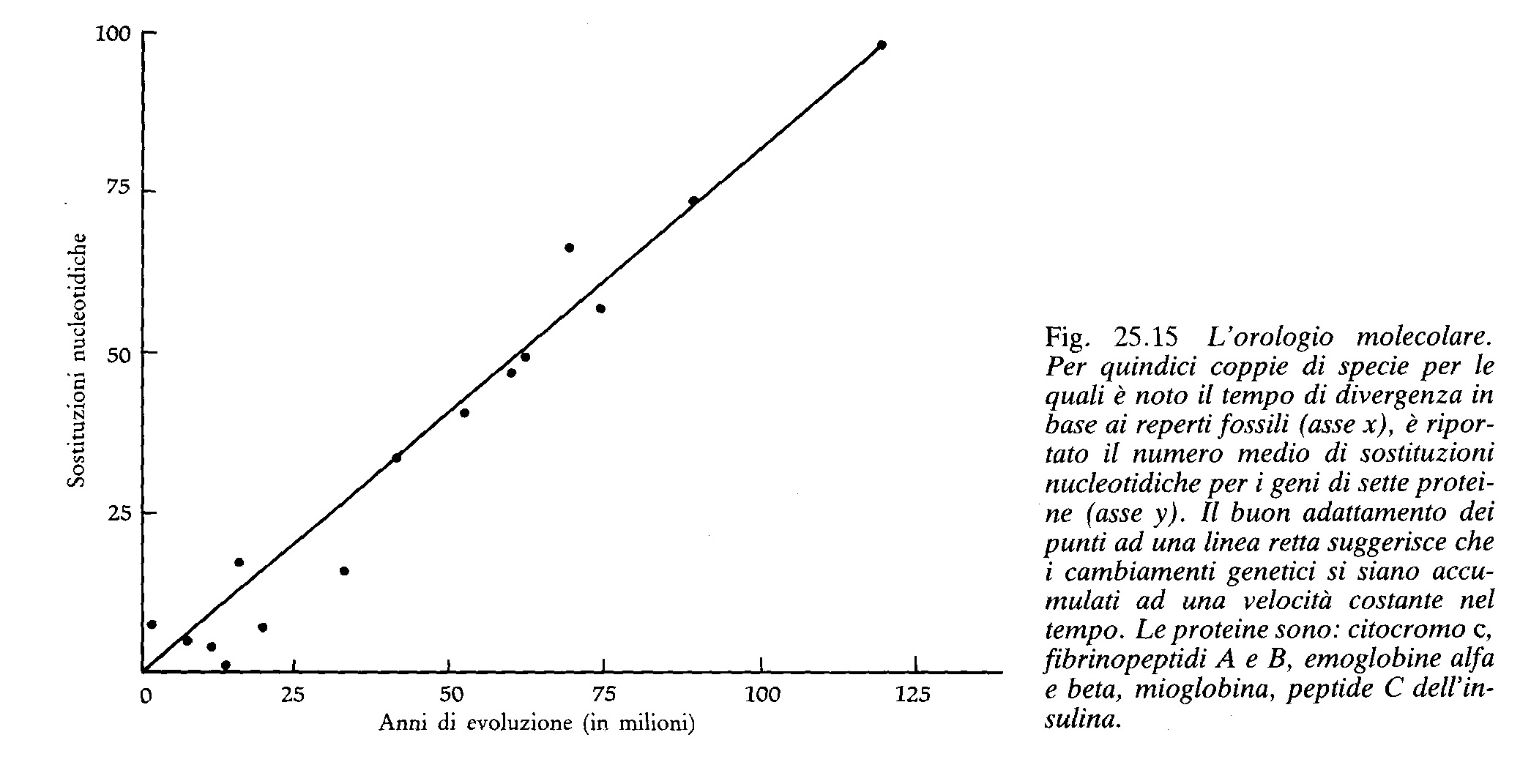
Questa velocità di evoluzione è sufficientemente costante perché possa essere usata come orologio? Consideriamo il citocromo c, un componente della catena respiratoria dei mitocondri. Il citocromo c dei vertebrati è formato da una catena di 104 aminoacidi; ad esempio, il numero medio di differenze aminoacidiche fra il genere Equus (cavalli e asini) e gli altri mammiferi è 5,1. Dal momento che i principali gruppi di mammiferi si separarono circa 90 milioni di anni fa, il tasso di sostituzione aminoacidica sarà 90/5,1 cioè una sostituzione ogni 17,6 milioni di anni. Possiamo ora saggiare se questo tasso è approssimativamente costante: dal momento che l'ultimo progenitore comune a rettili e mammiferi risale a 300 milioni di anni fa, la differenza media fra questi due gruppi dovrebbe essere di 300/17,6 = 17 aminoacidi; la differenza misurata, 14,8, è ragionevolmente simile a questo valore. Facendo affidamento su questo orologio possiamo datare gli eventi evolutivi per i quali mancano le evidenze fossili. Ad esempio i citocromi c delle piante e degli animali differiscono, in media, di 45,0 aminoacidi. Da questo valore si può stimare che la divergenza dall'antenato comune risale a 800 milioni di anni fa (45 cambiamenti x 17,6 milioni di anni ogni cambiamento = 792 milioni di anni).
Come vedremo nel capitolo 27 questa stima è ragionevole anche se molti paleontologi ritengono che tale evento si sia verificato ancora prima.
La validità dell'orologio molecolare è stata controllata per molte proteine ed è risultato che questo è un metodo piuttosto buono. Il tasso di cambiamento varia da una proteina all'altra ma per ciascuna proteina tende ad essere relativamente costante. L'orologio non è perfetto (un suo autorevole sostenitore dice che è un “orologio trasandato”) ma è assai più esatto di quanto molti darwnisti avrebbero potuto prevedere. Quale potrebbe essere l'origine di questa velocità costante? Se la maggioranza dei cambiamenti evolutivi avvenisse sotto il controllo della selezione naturale non ci si attenderebbe una velocità costante. La forza della selezione varia in base ai cambiamenti climatici, allo spostamento dei continenti, alla formazione di nuovi mari, all'evoluzione di nuovi predatori e ad altri fattori di intensità variabile nel tempo. In realtà sarebbe molto difficile immaginare una velocità di sostituzione costante nel tempo se l'orologio molecolare non funzionasse sullo stesso principio di quello radioattivo e cioè in modo puramente casuale. I neutralisti sostengono che la selezione naturale non casuale può essere responsabile solamente di una piccola parte dei cambiamenti aminoacidici; essi sostengono che, se l'orologio molecolare è esatto, l'evoluzione deve essere in gran parte scandita da cambiamenti neutrali.
Contro l'uso dell'orologio molecolare quale prova della neutralità selettiva della maggior parte dei cambiamenti selettivi i darwinisti portano due argomenti potenzialmente validi. Innanzitutto la selezione naturale, considerata su lunghi periodi, dovrebbe produrre una velocità di sostituzione costante anche se vi sono fluttuazioni temporanee intense. Ad esempio il Vesuvio erutta in modo imprevedibile e catastrofico dopodiché rimane quiescente per millenni (una variabilità, questa, molto maggiore di quella della selezione naturale). Tuttavia, considerando periodi di milioni di anni, si può calcolare un ritmo di eruzione pressoché costante.
In secondo luogo vi sono molte “date” molecolari importanti che non risultano essere in accordo con le informazioni paleontologiche più esatte. Dati molecolari di diversa origine forniscono un tempo di divergenza fra l'uomo e le grandi scimmie di circa 5 milioni di anni. I più antichi australopitecini identificati con certezza risalgono a 4 milioni di anni fa. Dal momento che gli australopitecini rappresentano già uno stadio avanzato nella via che conduce all'uomo (ad esempio avevano una stazione completamente eretta), la maggioranza degli antropologi ritiene che non possano essersi evoluti dall'antenato quadrupede comune alle scimmie e all'uomo in un solo milione di anni (esistono però cambiamenti evolutivi molto rapidi.).
La quota di variabilità nelle popolazioni naturali
Quando la selezione naturale agisce in una direzione definita, essa consuma variabilità in quanto tende a conservare un solo allele per ogni locus e a eliminare gli altri. Esistono molti processi che mantengono la variabilità, anche in presenza di selezione naturale: ad esempio una maggiore sopravvivenza degli eterozigoti oppure una pressione di selezione che subisce oscillazioni stagionali. Nel prossimo capitolo tratteremo questo argomento in dettaglio; per il momento diremo solamente che quegli evoluzionisti che fanno uso della matematica hanno elaborato modelli per prevedere la quantità massima di variabilità che si può mantenere in una popolazione quando l'unico agente del cambiamento evolutivo è la selezione naturale. Si è constatato che molte popolazioni conservano molta più variabilità di quella consentita da questi modelli basati sulla selezione, ma non si è accertato se ciò è dovuto al fatto che i modelli sono carenti oppure se la variabilità è realmente neutrale e non può essere eliminata dalla selezione.
Per queste ragioni i darwinisti replicano spesso che l'evoluzione non consiste nel mantenimento di geni invisibili nelle popolazioni. Sono i cambiamenti genetici significativi quelli che vengono tradotti in forme, funzioni e comportamenti che hanno rilevanza per gli organismi e, per quanto ne sappiamo, il successo o il fallimento di queste proprietà importanti è determinato dalla selezione naturale. I neutralisti possono essere d'accordo sul fatto che l'evoluzione della forma esterna e del comportamento sia determinata dalla selezione, ma sostengono che i geni che controllano queste proprietà significative costituiscono solo una piccola percentuale della variabilità di un pool genetico mentre la restante parte è neutrale in senso stretto oppure controllata dalla pressione di mutazione. Se è così la variabilità darwiniana (cioè quella che può essere “vista” dalla selezione naturale) è responsabile solamente di una piccola parte delle frequenze geniche che cambiano nel tempo. Chi può risolvere questa disputa? Quest'argomento dovrebbe essere affrontato con metodi empirici, ma per ora non disponiamo di mezzi per misurare quante delle modificazioni delle frequenze geniche siano adattative e quante neutrali.
La selezione naturale modella l'organismo ideale?
468 Tutte le idee più raffinate possono essere alterate e peggiorate. Anche l'idea della selezione ne è stata investita, in quanto alcuni suoi sostenitori troppo accesi le hanno attribuito lo stesso ruolo che i biologi settecenteschi attribuivano a Dio. Questi biologici pre-darwiniani sostenevano che la potenza di Dio si manifesta nella perfezione degli organismi -cioè nella superiorità della loro costruzione in senso ingegneristico e nel perfetto adattamento di questa all'ambiente. Alcuni biologi evoluzionisti moderni, sulla base di estrapolazioni troppo ardite sul mimetismo degli insetti-stecco o sulla superba costruzione dell'ala del gabbiano, hanno riesumato questo punto di vista sostituendo la selezione a Dio e sostenendo che qualsiasi sfumatura della forma organica ed ogni sottile differenza fra specie affini deve essere stata modellata dalla selezione naturale in quanto rappresenta la struttura ideale. Noi respingiamo un determinismo così rigoroso per diverse ragioni; la selezione non può ottimizzare tutte le singole parti di un organismo in quanto queste devono condividere le risorse metaboliche limitate del corpo. Se, ad esempio, ammettiamo che le immense corna dell'alce irlandese avevano un alto valore adattativo per le esibizioni sessuali (fig. 25.13) e che i maschi con le corna più grandi avevano un maggiore successo riproduttivo, perché esse non sono diventate ancora più grandi? Ad un certo punto il vantaggio di possedere corna grandi per scacciare gli altri maschi deve essere stato controbilanciato dal peso di queste strutture così ingombranti e dal costo metabolico richiesto per la rigenerazione annuale. La selezione lavora sull'intero organismo e non sulle singole parti e il disegno ottimale si ottiene solo dall'equilibrio fra le richieste contrastanti delle diverse parti.
Se le parti non sono ottimizzate cosa ne è degli organismi intesi come unità? La selezione produce sempre il migliore disegno costruttivo per un organismo nel suo ambiente? Vi sono biologi che sostengono questo punto di vista; la selezione, però, non è un principio che tende alla perfezione bensì un principio che tende solamente al miglioramento. La selezione non ha il compito cosmico di raggiungere un obiettivo prefissato e neppure realizza i progetti di un ingegnere. Se i geni di un organismo danno a questo un maggiore successo nel proprio ambiente, la selezione aumenta la sua fitness riproduttiva dopodiché si arresta finché non si originano nuove combinazioni genetiche ancor più favorevoli. Se gli ambienti rimanessero inalterati e le specie vi vivessero per innumerevoli milioni di anni, alla fine questo processo capriccioso potrebbe portare alla forma ottimale. Viceversa gli ambienti fluttuano perennemente, obbligando le popolazioni a seguire un ideale sempre diverso e non consentendo loro di avvicinarsi alla forma ideale.
Un altro fattore che si oppone all'idea della forma perfetta è rappresentato dall'incongruenza esistente fra il raggiungimento di una situazione ottimale per la popolazione e il fatto che la selezione agisce sui singoli individui. La selezione naturale è una lotta fra gli individui per raggiungere un maggior successo riproduttivo. Spesso potremmo individuare una strategia ottimale per il bene di una specie; se tutti i membri seguissero le regole ciascuno ne trarrebbe beneficio. Viceversa l'unica regola della selezione è il vantaggio individuale e ciò che sarebbe ottimale per la specie viene sacrificato a favore di una strategia stabile che tenga conto dei diversi modi di raggiungere il successo individuale.
Il biologo inglese John Maynard Smith ha fornito un complesso esempio teorico; supponiamo che in una specie esistano due strategie per procurarsi un partner, e che ciascun membro possa attuarne una sola delle due. I due gruppi di individui sono stati bizzarramente indicati come “falchi” e “colombe”: i falchi combattono fino alla fine, senza arrendersi a meno che siano gravemente feriti; le colombe, invece, si affrontano gonfiando le penne e minacciandosi ma non si colpiscono: la lotta termina quando uno dei due contendenti prevale sull'altro e l'unico prezzo pagato dal perdente è lo spreco di tempo e di energie. Quando un falco incontra una colomba quest'ultima batte in ritirata dopo il primo attacco e salva la pelle ma perde la possibilità di riprodursi.
Una popolazione composta da soli falchi si estingue rapidamente in quanto questi si uccidono fra loro. Una popolazione composta da sole colombe, invece, rappresenta la soluzione ottimale per il bene della specie. Il vincitore di uno scontro fra colombe ottiene il grande vantaggio dell'accoppiamento mentre per il perdente vi sono solo piccoli sacrifici. Ogni colomba vince circa la metà dei suoi combattimenti e la media fra i grandi vantaggi delle vittorie e le piccole perdite delle sconfitte rappresenta un vantaggio per tutti. Tutto ciò sarebbe molto bello se si eccettua un particolare. Se, per una mutazione, in questa popolazione nasce un falco, all'inizio esso incontrerà solamente colombe e, dal momento che vincerà tutti i combattimenti, troverà una forte selezione a suo favore. Nelle generazioni successive i falchi aumenteranno rapidamente ed inizieranno a scontrarsi tra loro; ciò controbilancerà i vantaggi ottenuti negli scontri con le colombe. Maynard Smith ha dimostrato che le popolazioni stabili sono composte in parte da falchi e in parte da colombe. Tutti gli individui staranno abbastanza bene perché i falchi incontreranno le colombe con una frequenza sufficiente ad annullare gli svantaggi degli incontri con altri falchi, mentre le colombe, pur perdendo in tutti gli scontri con i falchi, saranno avvantaggiate dal fatto di non essere ferite negli incontri con altre colombe. Il fatto più importante è che questa popolazione mista, pur essendo stabile, dà al singolo individuo un vantaggio minore rispetto a quella composta da sole colombe.
La popolazione di colombe rappresenta la miglior soluzione per il bene della specie. Perché non si evolve? Lo farebbe se la selezione operasse su interi gruppi a vantaggio della collettività; viceversa essa opera sui singoli individui e a loro unico vantaggio. Il falco mutante nella popolazione di colombe ottiene un enorme vantaggio selettivo anche se, complessivamente, riduce la fitness della popolazione. Dal momento che le colombe non possono arrestare il processo della mutazione genetica e che la selezione opera solamente sugli individui, la popolazione mista si può sviluppare anche se non rappresenta la soluzione ideale per la specie in toto.
L'idea che, grazie alla selezione naturale, gli organismi raggiungano una forma ottimale è smentita anche dall'esistenza di molte soluzioni di uno stesso problema: ad esempio gli elefanti africani hanno orecchie lunghe mentre quelli indiani hanno orecchie corte. I sostenitori dell'ottimizzazione dovrebbero concludere che, dal momento che la selezione produce sempre il meglio, deve esservi qualcosa nell'ambiente che in un luogo favorisce le orecchie lunghe e nell'altro le orecchie corte. Viceversa la maggioranza dei problemi ha molte soluzioni valide. La scelta di una di queste dipende dai casi della storia o dal fatto che, nelle prime tappe evolutive, sono state imboccate casualmente determinate direzioni. Quando una popolazione si è avviata in una di queste direzioni può rimanervi incanalata, anche se sarebbero potute esistere soluzioni migliori. Quando una nazione costruisce tutta la sua rete ferroviaria con un certo scartamento e tutto il materiale rotabile si adatta a questa misura, anche se qualche ingegnere dimostra che si otterrebbero risultati migliori con uno scartamento più ampio vi sarebbero molte difficoltà per cambiare la distanza fra tutte le rotaie. E possibile che le orecchie piccole rappresentino la soluzione migliore sia in Africa che in India, ma anche quelle grandi funzionano bene e fra gli elefanti indiani e quelli africani non c'è competizione. Quindi, se questo carattere è controllato dalla selezione, le soluzioni sono adattative: non è affatto necessario che siano quelle ottimali.
Il genetista americano Sewall Wright esprime l'idea delle soluzioni multiple con il concetto di paesaggio adattativo. Egli rappresenta l'universo delle soluzioni potenziali come un terreno con picchi e vallate: tutti i picchi sono adattativi ed alcuni di essi sono più alti di altri. Per sopravvivere una popolazione si deve trovare su un picco ma non ha bisogno di scalare quello più alto. Il caso e le circostanze storiche determinano il picco sul quale un popolazione tende a salire, ma l'ascensione è risultato della selezione.
La selezione naturale regola tutta la storia della vita?
469 Se si esclude la selezione naturale non è stata scoperta nessuna altra forza che possa essere l'agente che dirige l'evoluzione; è possibile che essa abbia modellato tutta la storia della vita? Possiamo confidare nel fatto che essa ha governato la storia organica per più di 3 miliardi di anni, dal mondo delle alghe e dei batteri all'avvento dell'Homo sapiens, mentre è possibile che sia estranea agli eventi primordiali e a quelli più recenti.
E probabile che l' “evoluzione” chimica che ha preceduto l'aggregazione primordiale delle forme organiche non sia avvenuta mediante processi paragonabili alla selezione naturale. Probabilmente buona parte di questa evoluzione è avvenuta mediante la produzione automatica di composti complessi in determinate condizioni fisiche (ad esempio la formazione degli aminoacidi nell'atmosfera primitiva per effetto delle scariche elettriche) e l'aggregazione di questi composti in determinate condizioni termodinamiche. La selezione naturale non ha potuto iniziare ad agire fino a che l'apparato chimico non ha sviluppato la capacità di commettere errori rari ed ereditabili durante la propria replicazione.
In termini geologici cinquantamila anni sono un istante - pochi secondi prima della mezzanotte se paragoniamo la storia della vita ad una giornata. Eppure in questo breve periodo la superficie terrestre è stata modificata con una rapidità che non è stata raggiunta neppure dalle più grandi catastrofi geologiche. Nel bene e nel male tutto ciò è opera di una sola specie, Homo sapiens, che lo ha realizzato soprattutto negli ultimi 300 anni. La selezione naturale ci ha formati ed ancora agisce su di noi come sulle altre specie, ma la sua forza è nulla a confronto dei cambiamenti delle nostre capacità provocati dall'evoluzione culturale - la trasmissione della conoscenza, delle tecniche e del comportamento realizzata tramite l'insegnamento e la tradizione. Non vi sono prove che indichino che la mente umana sia cambiata geneticamente dai tempi dell'antico Egitto o dell'uomo di Cro-Magnon, 50 000 anni fa. Tuttavia abbiamo accumulato delle spaventose capacità grazie alla nostra capacità di trasmettere l'informazione appresa mediante la cultura. L'importanza della selezione naturale è divenuta trascurabili per quanto riguarda la modificazione della nostra specie poiché essa deve operare secondo il modo darwiniano che è estremamente lento il metodo indiretto della variazione casuale e della conservazione dei varianti favoriti. Viceversa l'evoluzione culturale non è darwiniana. Essa è molto rapida poiché procede secondo il modo lamarckiano grazie all'ereditarietà dei caratteri acquisiti noi inventiamo una bomba nucleare e trasmettiamo il segreto della sua costruzione alle generazioni future. L'evoluzione della mente umana ha creato un'enorme discontinuità nella storia della vita in quanto ha dato inizio al primo processo naturale che può operare in modo lamarckiano. Questa constatazione non annulla in alcun modo la validità dell'affermazione secondo cui la selezione naturale ha diretto pressoché tutta l'evoluzione organica, ma vuole essere solamente un avvertimento del fatto che a questa si è aggiunto qualcosa di profondamente nuovo: un processo che ha soppiantato la selezione stessa e le ha conferito un ruolo subordinato nella determinazione della nostra storia futura.
Epilogo 470
Il più abile amico e sostenitore di Darwin, Thomas Henry Huxley, si sentiva molto deluso per non avere scoperto personalmente un principio così semplice e potente come è la selezione naturale. Egli si proclamò “estremamente stupido” per aver perduto una simile occasione. Forse nella storia della scienza non c'è stato nessun altro concetto altrettanto semplice nella costruzione - pochi fatti evidenti ed una deduzione ovvia - e contemporaneamente ostico alla comprensione generale. Gli ostacoli che esso ha incontrato nascono dalla scarsa propensione di molte persone ad abbandonare idee antiquate ma comode che non si conciliano con la selezione naturale - soprattutto la nozione secondo cui l'evoluzione deve implicare un progresso immanente ed ha un suo scopo e la convinzione che l'Homo sapiens occupi un posto speciale e prefissato nell'economia della natura. Eppure, malgrado il suo potere nel corso dell'evoluzione, la selezione naturale non è onnipotente. Gli organismi non sono macchine ottimali progettate dalla selezione naturale per essere i migliori oggetti concepibili per i loro ambienti. Essi sono i prodotti di una storia di miliardi di anni che li ha obbligati a variare e a modificarsi in un numero di modi limitato.
Sommario 471
1. Le migliori prove dell'evoluzione sono quelle anomalie e imperfezioni della forma che riflettono la storia trascorsa. Ad esempio: se le balene con fanoni non si fossero evolute da antenati con denti normali perché i loro embrioni avrebbero dovuto sviluppare i denti per poi riassorbirli senza farne uso? Perché il braccio di un uomo, la pinna di una balena, l'ala di un pipistrello o la zampa di un cavallo - strutture con funzioni estremamente diverse - dovrebbero essere costruite con le stesse ossa se queste non fossero un retaggio dell'evoluzione da un antenato comune?
2. La teoria di Darwin della selezione naturale è un meccanismo in due tappe che spiega le modificazioni evolutive: prima la variazione casuale si comporta come materiale grezzo; dopo la selezione naturale conserva in modo differenziale quelle porzioni dell'intervallo di variazione che offrono un maggior adattamento all'ambiente locale. Ad esempio se per gli individui è vantaggioso essere più piccoli lo scostamento dalle dimensioni ancestrali è ugualmente probabile in tutte le direzioni, ma la selezione naturale definisce la direzione del cambiamento evolutivo attribuendo un maggior successo riproduttivo agli individui più piccoli della media.
3. Il principio darwiniano della selezione naturale ha molte implicazioni che non si conciliano con le speranze umane e con le tradizioni culturali occidentali. Esso nega qualsiasi tendenza intrinseca al progresso che debba portare necessariamente all'evoluzione dell'uomo ma parla soltanto di adattamento ad ambienti locali mutevoli. Esso non contiene alcuna affermazione di principio circa il «bene della specie» o «l'armonia degli ecosistemi», sostenendo che l'ordine e l'armonia della natura si originano come sottoprodotti della selezione che attribuisce ai singoli individui un maggior successo riproduttivo.
4. Le azioni altruistiche (in cui un individuo si pone in una situazione di pericolo o si sacrifica per aiutare gli altri membri della specie) appaiono come una contraddizione della nozione darwiniana secondo cui la selezione naturale agisce unicamente mediante il successo riproduttivo. Il successo riproduttivo deve essere definito come un maggior contributo offerto con i propri geni alle generazioni future (il corpo in quanto tale non è ereditato). I parenti hanno in comune copie di geni altruistici. Se un individuo esegue azioni altruistiche a vantaggio di parenti sufficientemente stretti può esservi un vantaggio maggiore per i geni altruistici rispetto al caso in cui l'individuo ignora i suoi parenti ed agisce unicamente a proprio vantaggio. Questa teoria, detta della kin selection, rappresenta un corollario del darwinismo classico e può spiegare fenomeni quali la «collaborazione al nido» (uccelli che rinunciano alla propria riproduzione e si prendono cura dei nidi dei propri parenti) e la sterilità delle api e delle formiche operaie.
5. L'evoluzione all'interno delle popolazioni è definita come il cambiamento delle frequenze dei geni nel corso del tempo. La selezione naturale non è il solo meccanismo che può portare al cambiamento delle frequenze geniche. Nelle piccole popolazioni acquistano rilevanza due meccanismi di cambiamento casuale: il principio del fondatore, secondo cui nelle nuove popolazioni fondate da pochi individui mancano alcuni alleli della popolazione parentale e gli altri sono presenti con frequenze diverse; la deriva genetica, per cui alcuni geni sono eliminati ed altri aumentano di frequenza per effetto del caso e non del loro valore selettivo.
6. La terza posizione del codice genetico è ridondante - cioè spesso le mutazioni in questa posizione non modificano l'aminoacido codificato dalla tripletta - e pertanto sfugge alla selezione naturale (che agisce soltanto sui prodotti del codice genetico). Vi possono essere anche altre mutazioni neutrali, oppure così poco nocive che sono eliminate dalla selezione più lentamente di quanto esse si riformino a causa di nuovi eventi mutativi. La frequenza di queste mutazioni neutrali può aumentare ed esse possono prendere il sopravvento anche in popolazioni di grandi dimensioni.
7. Se il cambiamento evolutivo consiste nella fissazione di mutazioni neutrali la velocità delle modificazioni a livello genetico dovrebbe essere costante. Alcuni scienziati ritengono che questo «orologio molecolare» esista effettivamente (esso è stato tarato confrontando le modificazioni della struttura delle proteine in organismi differenti con i tempi di divergenza dedotti dalle testimonianze fossili). Altri sostengono che questo orologio è troppo imperfetto per registrare le mutazioni neutrali, oppure che la sua costanza non è in contraddizione con-la presenza di un controllo da parte della selezione naturale.
8. La selezione naturale non modella gli organismi ottimali. Essa migliora gli organismi in funzione dell'ambiente locale, ma non li rende perfetti. Il raggiungimento della perfezione è impedito dalle condizioni imposte dalla struttura e dall'esistenza di molte soluzioni valide per i problemi di adattamento (se gli organismi scelgono una soluzione valida ma non ottimale potranno insorgere problemi quando la necessità di specializzare gli adattamenti imporrà cambiamenti futuri).
9. La selezione naturale è stata la forza più importante che ha determinato i cambianienti durante la storia della vita, anche se non è intervenuta in tutti gli eventi. All'origine della vita essa ha potuto iniziare a funzionare solamente quando i composti chimici complessi svilupparono la capacità di produrre rari errori ereditari della replicazione. L'influsso della selezione naturale è stato scarso anche per quanto riguarda l'aumento di complessità della tecnologia e della società registrato negli ultimi 50 000 anni della storia umana. In questo intervallo di tempo le dimensioni del nostro cervello non sono aumentate e questo aumento di complessità e di raffinatezza riflette probabilmente un'evoluzione culturale e non un cambiamento biologico.